棉花枯萎病的发生与为害
棉花枯萎病的发生与为害
棉花枯萎病[
一、发生与蔓延
自1891年Atkinson首先在美国发现并报道枯萎病以后,陆续在其他产棉国和地区相继有发生和为害的报道。到21世纪初,棉花枯萎病的发病地区已遍及亚洲、非洲、北美洲、南美洲及欧洲等地,包括印度、巴基斯坦、缅甸、扎伊尔、埃塞俄比亚、加蓬、乌干达、乍得、南非、苏丹、坦桑尼亚、美国、阿根廷、巴西、秘鲁、乌拉圭、委内瑞拉、圣文森特岛、希腊、意大利、南斯拉夫、俄罗斯和澳大利亚等,几乎已遍及世界各植棉国家。
随着美国棉花品种种子的引进,枯萎病传入我国。1931年冯肇传首先在华北地区发现棉花枯萎病;1934年黄方仁报告棉花枯萎病在江苏南通发生为害;1936年沈其益报道在南京、上海发现棉花枯萎病。以后随着棉花种子的繁殖、调运和推广,病区逐年扩大,为害也日趋严重。
1949年前,棉花枯萎病已扩展到7个省、市、自治区,其中,陕西泾惠灌区由兴平至咸阳一线和泾灌区的局部棉田,山西曲沃、临汾一线、四川涪江两岸的射洪、三台等县,江苏省的南京、南通、启东和上海市,辽河流域的盖州、营口,河南弈川,安徽萧县,浙江慈溪,云南宾川,河北
(单位:发病县数)省(区、市)南方棉区19631982省(区、市)北方棉区19631982江苏349河南580四川1347山东172湖北643河北069安徽135陕西1843浙江226山西1437湖南020辽宁715江西016新疆*110广东02甘肃-4云南22北京010上海210天津-3总计29250总计46343
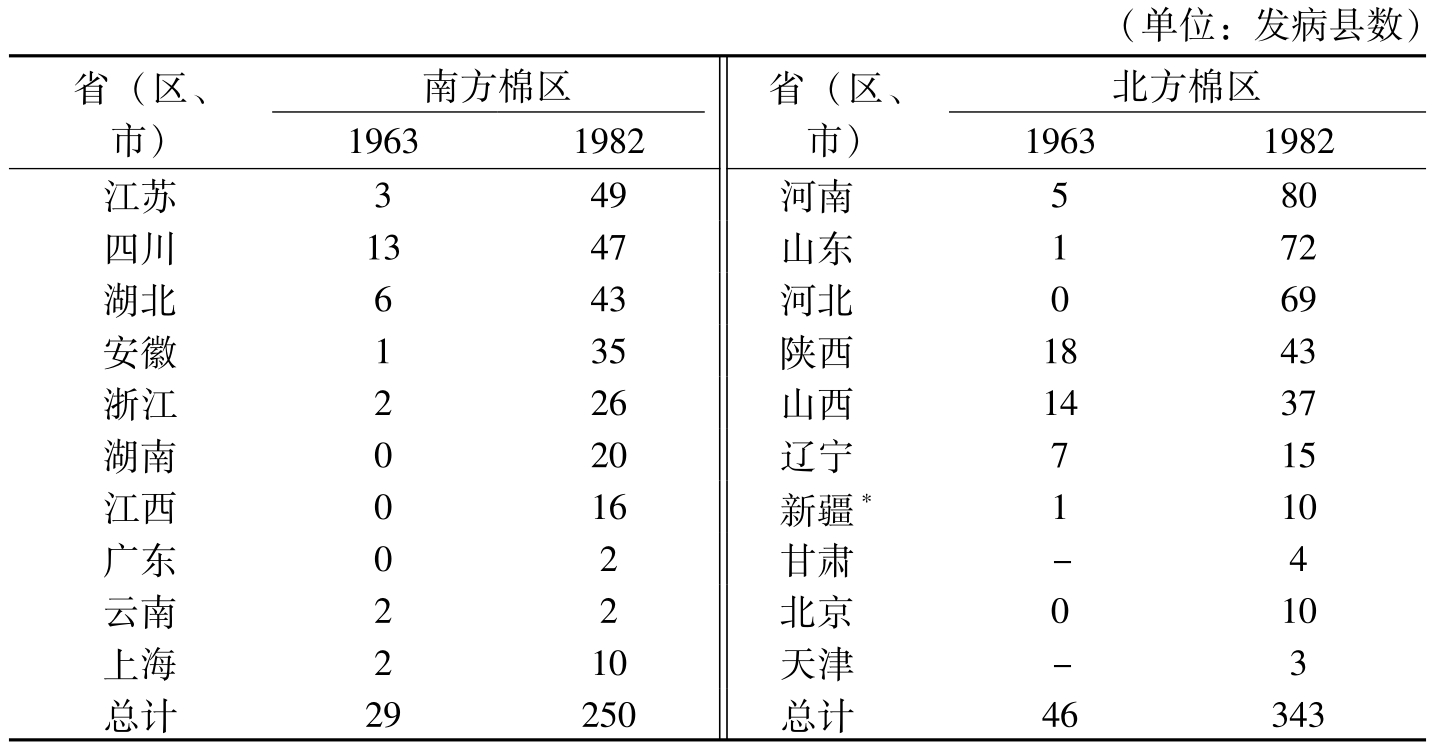
表1-1 棉花枯萎病在各棉区扩展蔓延概况比较(马存等,2007)
20世纪80年代初我国大力推广抗枯萎病棉花品种,而抗病品种的棉种也可能携带少量枯萎病菌,使枯萎病发病面积进一步扩大,但是由于抗病品种在重病田发病也很轻,到80年代末枯萎病的严重为害已得到控制。从30年代枯萎病传入到被控制,马存等(2007)把我国棉花枯萎病的发生分为以下4个阶段:第一阶段,20世纪30年代到新中国成立为传入阶
二、病害症状
棉花枯萎病病原菌从根部侵入、系统侵染为害棉株,因此,从苗期到成株期的各个生育阶段,均可表现病症。
概括起来,枯萎病的症状类型有:黄化型、黄色网纹型、紫红型、青枯型、半边黄型、皱缩型、枯斑型和光秆型。其中,前面4个症状多见于苗期阶段;黄色网纹型、半边黄型、皱缩型是枯萎病病株的典型症状类型。病株的症状是受病株一系列病理生理变化的外部反应,与病原菌致病力的强弱也有一定的关系。此外,症状表现往往因品种和自然环境的不同而有差异,如抗病品种中的感病株多呈青枯型、黄化型。而同期的感病品种则有多种症状型,气温低时为紫红型,雨后转晴时呈青枯型,严重的呈皱缩型、枯斑型,再严重的呈光秆型。在生产上,往往几种症状型混生于一株上,尤其是枯、黄萎病混合发生田,混生的症状类型更为普遍。
在枯、黄萎病混生地区,两病可以同时发生在一株棉花上,叫做同株混生型,有的以枯萎病症状为主,有的以黄萎病症状为主,使症状表现更为复杂,调查时需注意加以区分。兹将两病发病症状比较如表1-2所示。
项目枯萎病黄萎病株型植株节间缩短弯曲、有时顶端枯死一般植株不矮缩,顶端不枯死枝条有半边枯萎,半边无症状下部发病叶片可长出新枝叶叶片顶端叶片一般先显症下部叶片先显症,逐渐向上发展
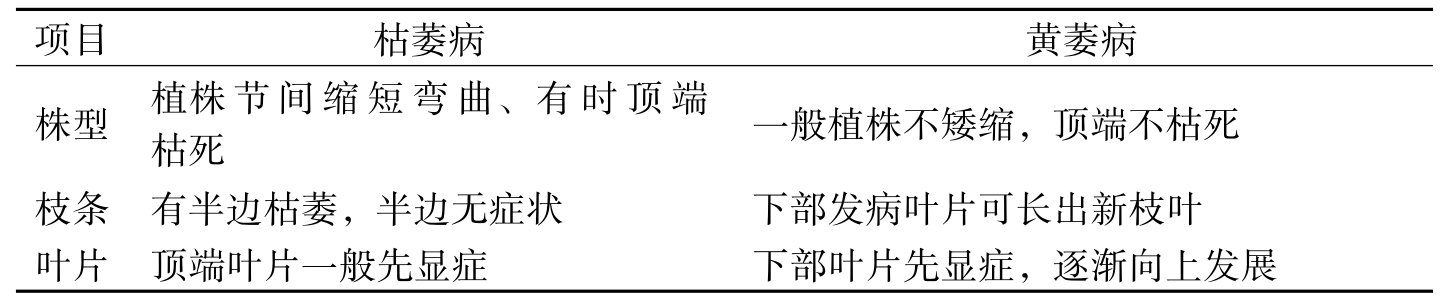
表1-2 棉花枯萎病、黄萎病的症状比较
项目枯萎病黄萎病叶脉叶脉常发黄色,呈明显的黄色网纹叶脉绿色、脉间叶肉及叶绿变黄色呈斑块状叶形常增厚、皱缩,深绿色,叶缘向下弯曲大小形状无异,叶缘向上卷曲维管束木质部变褐色明显,色深褐木质部变色,色淡褐

表1-2 棉花枯萎病、黄萎病的症状比较(续)-1
在田间普查诊断棉花枯萎病时,除了观察病株外部症状外,必要时应剖开茎秆检查维管束变色情况。感病严重植株,从茎秆到枝条甚至叶柄,内部维管束全部变色。一般情况下,枯萎病株茎秆内维管束呈褐色或黑色条纹。调查时剖开茎秆或掰下空枝、叶柄,检查维管束是否变色,这是田间识别枯萎病的可靠方法,也是区别枯萎病与红(黄)叶茎枯病,排除旱害、碱害、缺肥、蚜害、药害等原因引起类似症状的重要依据。此外,由于枯萎病维管束变色的深浅不是绝对的,有时黄萎病重病株比枯萎病轻株维管束变色还要深些,这就需要辅之以室内分离鉴定工作。
三、为害损失
枯萎病会影响棉花的正常生长发育,在生长前期,枯萎病可造成死苗,中后期则影响蕾铃生长发育直至脱落,导致棉花产量严重下降,纤维品质变劣。
关于枯萎病为害产量的损失,早期已有报道,Ware等(1934)调查研究认为,植株发病死亡率10%应该作为一个经济阈值,如高出这个数字,将会给产量带来负面影响。Chester(1946)估计,植株死亡率在60%之内,死亡率每增加5%,产量就要大约损失3%。20世纪80年代末,美国在全国范围内进行棉花枯萎病为害产量损失评估,1989年估计损失皮棉0.2%(Blasingame,1990)。也有报道重病年份损失皮棉1.05亿kg。姚耀文等(1963)研究结果指出,感枯萎病棉株的铃重(4.94g)、衣指(6.08g)、籽指(10.35g)和单株籽棉产量(60.91g)分别比健株减0.71g、0.59g、1.45g和31.75g,减少百分率分别为12.5%、8.8%、
棉株感染枯萎病后对棉花产量影响很大,病株的皮棉产量和籽棉产量均比健株极显著减产。具体表现在单株成铃数极显著减少;衣分显著降低;单铃重明显下降(表1-3和表1-4)。
病级皮棉产量(g/株)为0级的(%)籽棉产量(g/株)为0级的(%)衣分(%)单铃重(g)单株铃数(个)0级13.67aA100.0032.45aA100.0042.34aA4.09aA7.94aAⅠ级10.05bB73.5224.45bB75.3541.17bAB3.90abAB6.20bBⅡ级7.45cC54.5018.41cC56.7340.45bcB3.64bcBC4.79cCⅢ级6.21cC45.4315.48cC47.7040.04cB3.38cC4.49cCⅣ级3.04dD22.247.85dD24.1938.79dC2.88dD2.62dD

表1-3 棉株枯萎病对产量的影响(吴征彬等,2004)
病情分级单铃重(g)衣分(%)籽指(g)衣指(g)0级5.341.410.97.7Ⅰ级4.240.29.86.7Ⅱ级3.640.19.66.4Ⅲ级3.241.17.85.9Ⅳ级2.340.06.94.6
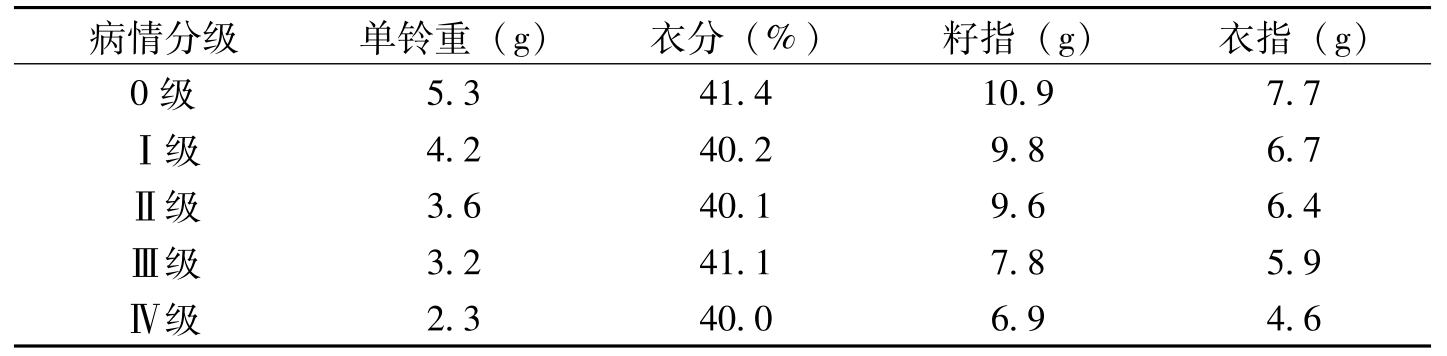
表1-4 棉花枯萎病对产量的影响(万英,2005)
吴征彬等(2004)对棉花枯萎病与棉花产量之间回归分析结果指出,当棉株枯萎病每提高1个级别,其单株皮棉产量的减产率为18.361%,单株籽棉产量减产率为17.927%,其衣分约下降0.824个百分点,单铃重约下降0.293g,单株结铃约减少1.235个。
棉株感染枯萎病后,其纤维长度等品质指标与健株相比都有显著降低。姚耀文等(1963)报道,感枯萎病棉株的纤维长度(30.1mm)和单强(3.69g)分别比健株减少2.0mm和0.64g。棉株的发病级别与纤维品
病级纤维长度(mm)整齐度(%)比强度(cN/tex)伸长率(%)马克隆值0级30.48aA85.95aA31.03aA8.14aA4.55aAⅠ级30.01bAB84.23bB29.75bAB7.62bB4.39aABⅡ级29.60bBC84.08bB29.53bB7.51bB4.35abABⅢ级29.11cCD83.91bB28.90bcBC7.51bB4.15bBⅣ级28.82cD83.80bB28.08cC7.50bB3.81cC
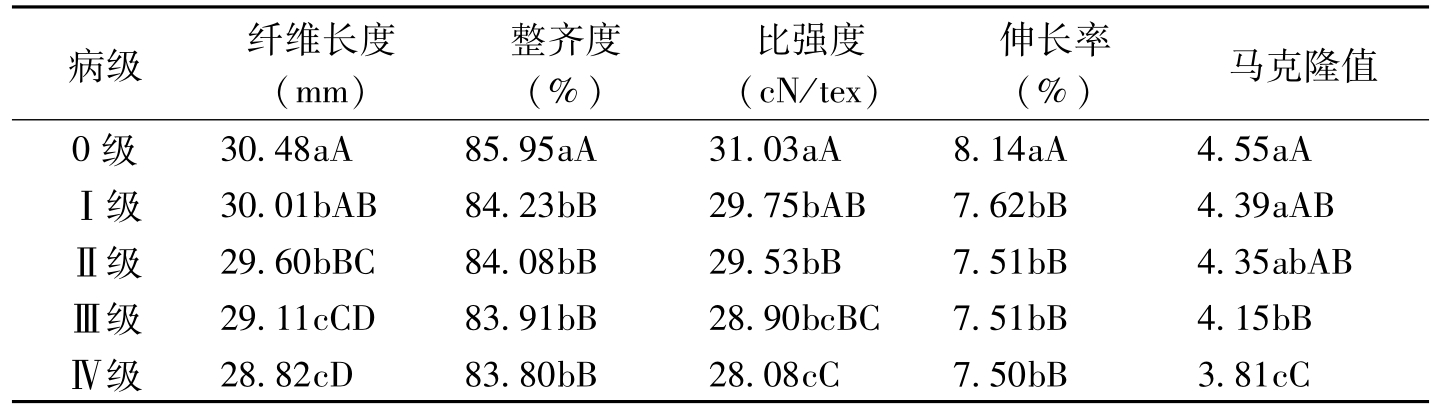
表1-5 棉花枯萎病对纤维品质的影响(吴征彬等,2004)
病级马克隆值比强度(cN/tex)纤维长度(mm)0级4.129.429.7Ⅰ级3.727.928.8Ⅱ级3.526.227.7Ⅲ级3.225.227.3Ⅳ级2.922.425.8
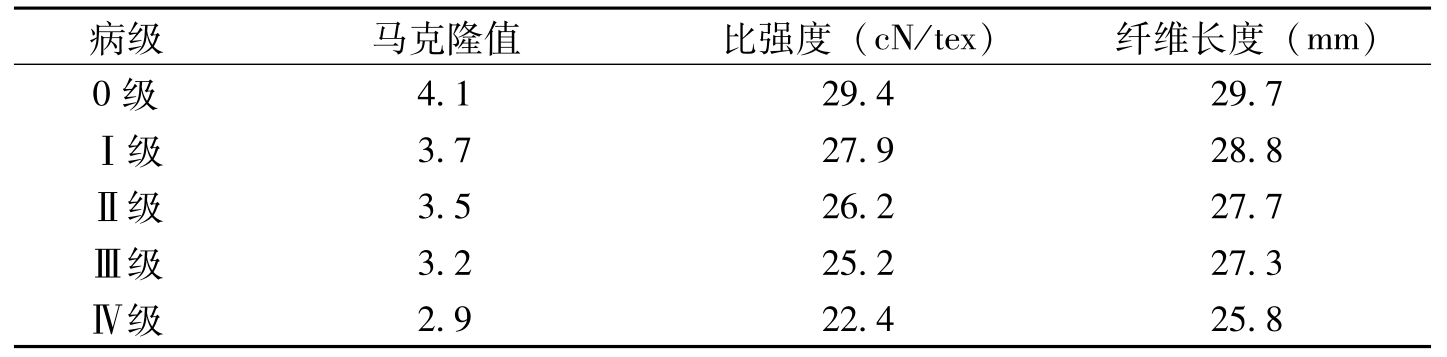
表1-6 棉花枯萎病对纤维品质的影响(万英,2005)
四、发病条件
棉花枯萎病的发生,除了与病原菌的种、生理小种或生理型的不同致病力等致病因素有关外,还与气候条件、土壤耕作栽培条件、品种和生育阶段等条件有关。
(一)与气象条件的关系
众多研究结果表明,温度是影响棉花枯萎病的重要因素之一。棉花枯萎病菌在培养基上生长的最适温度为18~25℃,最高为35℃,最低为5℃;而在田间,当土壤温度超过28℃时,棉花播种12天即可侵染发病,
在适温条件下,降水量是影响棉花枯萎病发生程度的一个重要因素。棉花枯萎病的发展情况还取决于发病期雨量的多少与分布。一般6~7月雨水多、分布均匀,则发病重;如果雨量少或降雨集中,则发病轻。沈万陆
(二)与土壤耕作栽培条件的关系
棉花生长发育除需要适宜的气候条件外,也需要良好的土壤耕作栽培条件,良好的土壤和耕作栽培条件可以促进棉花健壮生长,并能增进其对病害的抗病能力;反之则棉花生长不良,抗病性降低。
枯萎病菌在棉田定植以后,连作棉花年限愈长,土壤中病菌量积累愈多,病害就会愈严重。河南新乡县七里营公社调查(1977),连作2年棉田枯萎病发病率为4%~31.5%,死苗率为4.5%,连作3年的棉田发病率达36%~42%,死苗率为4%~9%;连作4年的棉田发病率高达58%~71%,死苗率达9%~12%。
棉花枯萎病菌在pH值2.8~9.0范围内的培养基上均能生长,说明其对酸碱度的适应性较广,但最适宜的pH值范围是在3.0~5.5。因此,枯萎病菌在田间引起棉花发病,对土壤酸碱度的适应范围较广,没有十分严格的界限。棉田地势低洼、排水不良,或者灌溉棉区,一般枯萎病发病较重。灌溉方式和灌水量都能影响发病,大水漫灌往往起到传播病菌的作用,并造成土壤含水量过高,不利于棉株生长而有利于病害的发展。
营养失调也是促成棉株感病的诱因。氮、磷是棉花不可缺少的营养,但偏施或重施氮肥,反而能助长病害的发展。原河南农学院(现河南农业大学)(1976)研究棉花营养与枯萎病发生的关系认为,单施氮肥较之单施磷肥或钾肥,发病率显著提高。氮磷钾配合适量施用,将有助于提高产量和控制病害发生。
不同类型肥料的施入也会改变局部土壤状况,进而影响棉花枯萎病的发生发展。简桂良等(1996)通过田间及盆栽试验研究认为,棉花枯萎病菌抑菌性土壤施入不同形式的有机肥后,抑菌性发生了变化。施入马粪后,抑菌性被削弱,枯萎病菌增殖加快,棉花枯萎病发病率与导菌土相当,而施入棉籽饼及豆饼之后,则抑菌性增强,抑菌效果增加,尖孢镰刀菌萎蔫专化型的增殖受到抑制,而非致病性镰刀菌及产荧光假单胞菌含量增加。Elad等(1985)认为,荧光假单胞菌与病原镰刀菌在根际和土壤中通过对根际碳素营养和铁离子的竞争,使病原菌由于缺乏营养及厚垣孢子萌发所必需的铁离子,从而无法萌发及增殖;而非致病性镰刀菌对根表侵染位点的竞争及组织的专性位点的定殖,从而使病原菌失去这些生态点,病害因而减轻。
不同育苗方式对棉花枯萎病有一定影响。阚画春等(2006)报道,营养钵育苗移栽的棉花比直播的发病轻,无土育苗移栽的棉花发病比营养钵育苗移栽的轻,地膜覆盖的棉花发病比无覆盖的轻(表1-7)。
直播营养钵育苗移栽无土育苗移栽直播地膜覆盖营养钵覆膜移栽无土育苗覆膜移栽发病株率(%)32.3911.253.3533.336.991.46病指26.564.382.0619.493.600.49
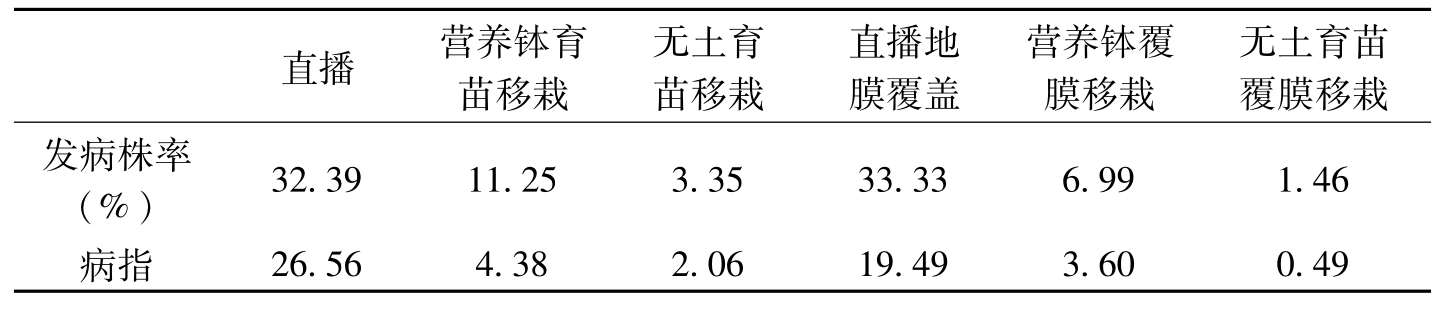
表1-7 不同育苗方式与枯萎病发病关系
刘海艳等(2008)通过2005~2006年病情系统调查和气象因子分析,查明新疆地膜棉田枯萎病的发生和内地一样有两个明显的发病高峰,并且与气温、降雨有十分密切的关系,但在地膜覆盖植棉的情况下,其发病比内地明显提高,当连续5天的平均气温达14℃左右,田间开始陆续出现病苗,当连续5天的平均气温达20℃左右,进入迅速发病期,很快出现第一个发病高峰;当连续5天的平均气温达24℃以上,则进入病情减轻或高温隐症期;7月下旬或8月上旬随温度下降,病情又有所回升,进入第二个发病高峰。降雨与棉花枯萎病的发生也有一定的关系,在6月15日以前,累计降水量越大的年份,枯萎病发生也越严重。
(三)与棉花品种、生育期的关系
棉花不同的种或品种,对枯萎病的抗病性具有很大差异。一般亚洲棉对枯萎病抗病性较强,陆地棉次之,海岛棉较差。在陆地棉中各品种间对枯萎病的抗性差异也很显著。20世纪70年代前育成的52-128、陕棉4号、86-1号等品种抗病性很强,尤其是52-128已成为我国抗枯萎病育种的抗原。
棉花枯萎病的田间发病与棉株的不同生育阶段也有一定的关系。虽然在温室内人工接种棉苗,均可在子叶展平期或真叶期出现症状,尤以三叶期更易接种成功。但田间的症状出现期,棉枯萎病多在5月见症,蕾期(即5月底至6月份)为显症高峰。当然发病高峰与这段时间内的适宜温、湿度有关。中国农业科学院植物保护研究所(1974)进行的病圃分期播种试验,设4个播种期,从出苗到出现发病高峰,尽管分别经历29~55天时间,但都是在现蕾前后进入发病盛期,若现蕾期推后则发病高峰也顺延,发病高峰的出现不因早播而提前。马存等(1980)在河南调查发现,5月26日棉花处于3~4片真叶期,枯萎病指为16.5;至6月3日,棉花进入现蕾期,病指上升到43.4,进入发病高峰期。如果分别于3月20日、4月15日、5月15日和6月15日播种,检查结果,病指高峰值32.1、43.0、31.0和8.9都是在现蕾期。
综上所述,棉花枯萎病发生与为害的主要影响因子是气候、土壤与耕作栽培等环境条件,棉花品种和生育阶段对棉花枯萎病亦有重要影响。发病高峰期主要出现在棉花现蕾期,若在现蕾期土温在25℃左右且连续阴雨,将会造成枯萎病暴发,高温干旱则造成症状的隐蔽,发病较轻。育苗移栽、地膜覆盖及合理增加种植密度等科学的栽培管理可以减轻棉花枯萎病的为害。
(四)与棉田线虫的关系
线虫是一类体形细长、不分节、无色透明的无脊椎动物。在自然界中,线虫的种类仅次于昆虫,但数量比昆虫还要多。据估计线虫有50多万种之多。植物寄生线虫主要存在于土壤中,根据在植物上的寄生部位可分
据国内外研究报道,已知为害棉花的植物线虫属和种主要有根结线虫(
棉田植物寄生线虫和棉花枯萎病的发生有一定关系。Orlon(1910)指出,根结线虫的严重侵染,必然会破坏棉花对枯萎病的抗性。Martin等(1956)用南方根结线虫和枯萎菌联合接种,枯萎病发病率显著提高。Holdeman等(1954),用刺线虫(
线虫不仅可单独发生,直接造成棉花经济损失,还可与真菌、细菌和病毒病原协同发生,构成线虫—病害复合症,加重病害对棉花的为害。
线虫在病害复合症中的作用主要是直接为害棉花之后改变植株的生理状况,并造成寄主植物(棉花)机械损伤,成为病菌侵入寄主的孔口,使病害加重;枯萎病菌侵入木质部导管需经由根尖组织,线虫在棉花1~2片真叶以后主要为害须根和根尖,造成伤口,有利于病菌的侵入;线虫取食
线虫在病害复合症中,作为致病诱因,常使棉品种对枯萎病的感病性提高,线虫和病菌并存而协同发生,棉花枯萎病发生更加严重,Carber等(1953)指出,在有根结线虫存在时,棉花表现为高度感染枯萎病,但在没有根结线虫存在时,棉花则高抗枯萎病,表现了根结线虫对棉花感枯萎病的诱因作用。Holdeman等(1954)试验结果,单独接种枯萎病菌,抗性品种Coker 100不发生枯萎病;当刺线虫和枯萎病菌联合接种时,抗病品种的枯萎发病率为60%,感病品种的发病率为78%,证明不论是抗病品种,还是感病品种,刺线虫都大大地促进棉花枯萎病的发生。
王汝贤等(1998)通过田间及温室盆栽试验,证明供试的3类线虫(N
年份(1)接种物(2)发病率(%)播后天数303743检验(3)(43天)发病率(%)播后天数303743检验(4)(43天)1985N1+F54.295.2100F42.172.283.3N2+F22.233.980.0F19.032.477.8CK0001.2340.23134.757.977.427.950.681.48.317.452.510.321.256.90002.0330.6771986N1+F33.373.386.7F30.480.070.0N3+F13.333.373.3F20.040.073.3CK0000.68213.345.077.313.836.352.55.022.353.313.335.066.70002.5241.490
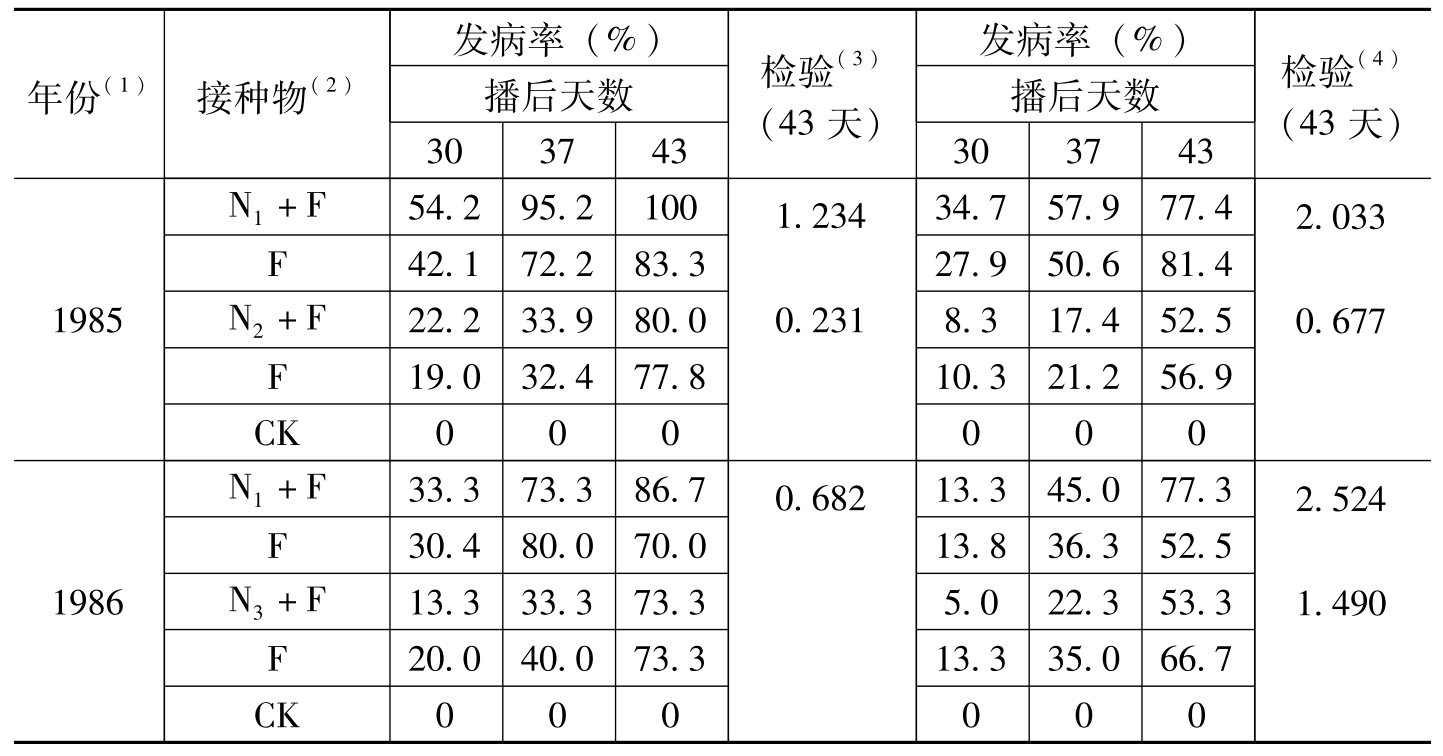
表1-8 不同类型线虫与棉花枯萎病菌的协同作用*
年份(1)接种物(2)发病率(%)播后天数303743检验(3)(43天)发病率(%)播后天数303743检验(4)(43天)1987N1+F22.244.472.2F16.729.252.2N3+F12.525.050.0F16.729.252.2CK0000.9830.51612.523.654.27.315.634.83.112.525.07.315.634.80002.6640.971
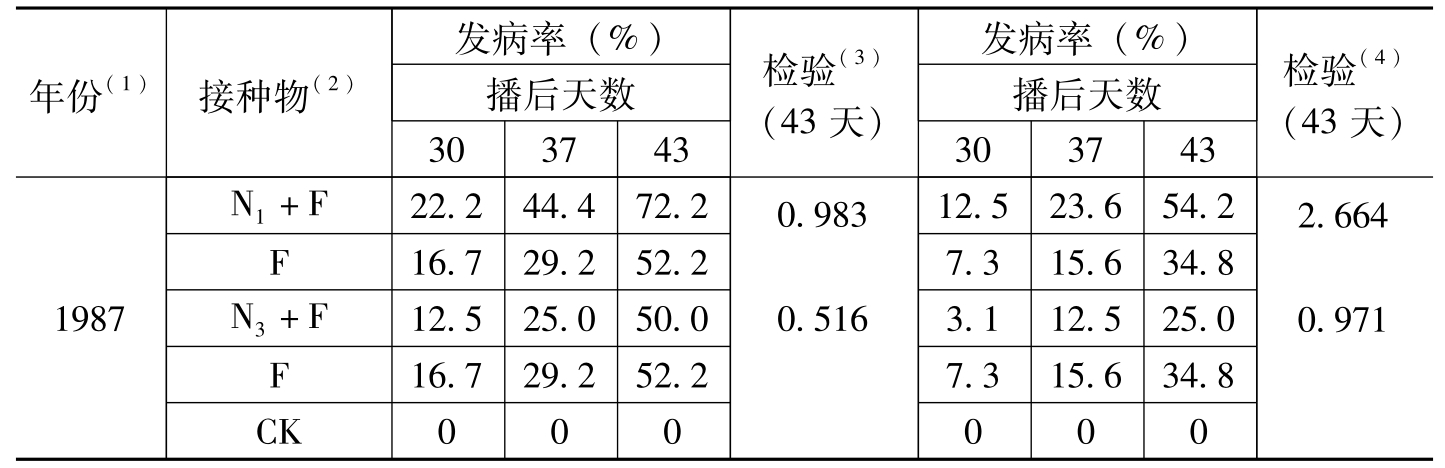
表1-8 不同类型线虫与棉花枯萎病菌的协同作用*(续)-1
邓先明等(1992,1993)报道,无论感病或抗病品种盆栽棉株单接肾形线虫(
品种日期(日/月)发病株率(%)病指ABA-BABA-B洞庭1号(感病品种)11/1119.316.28.08.04.53.518/1170.752.018.739.821.018.825/1189.857.232.672.028.843.22/1294.859.534.380.737.143.69/12100.070.729.390.845.545.3平均74.951.123.858.327.430.9川73-27(感病品种)11/116.706.71.601.618/1115.06.78.36.62.14.525/1125.08.316.711.63.38.32/1230.013.316.717.05.811.29/1238.316.721.625.49.216.2平均23.09.014.012.44.18.3
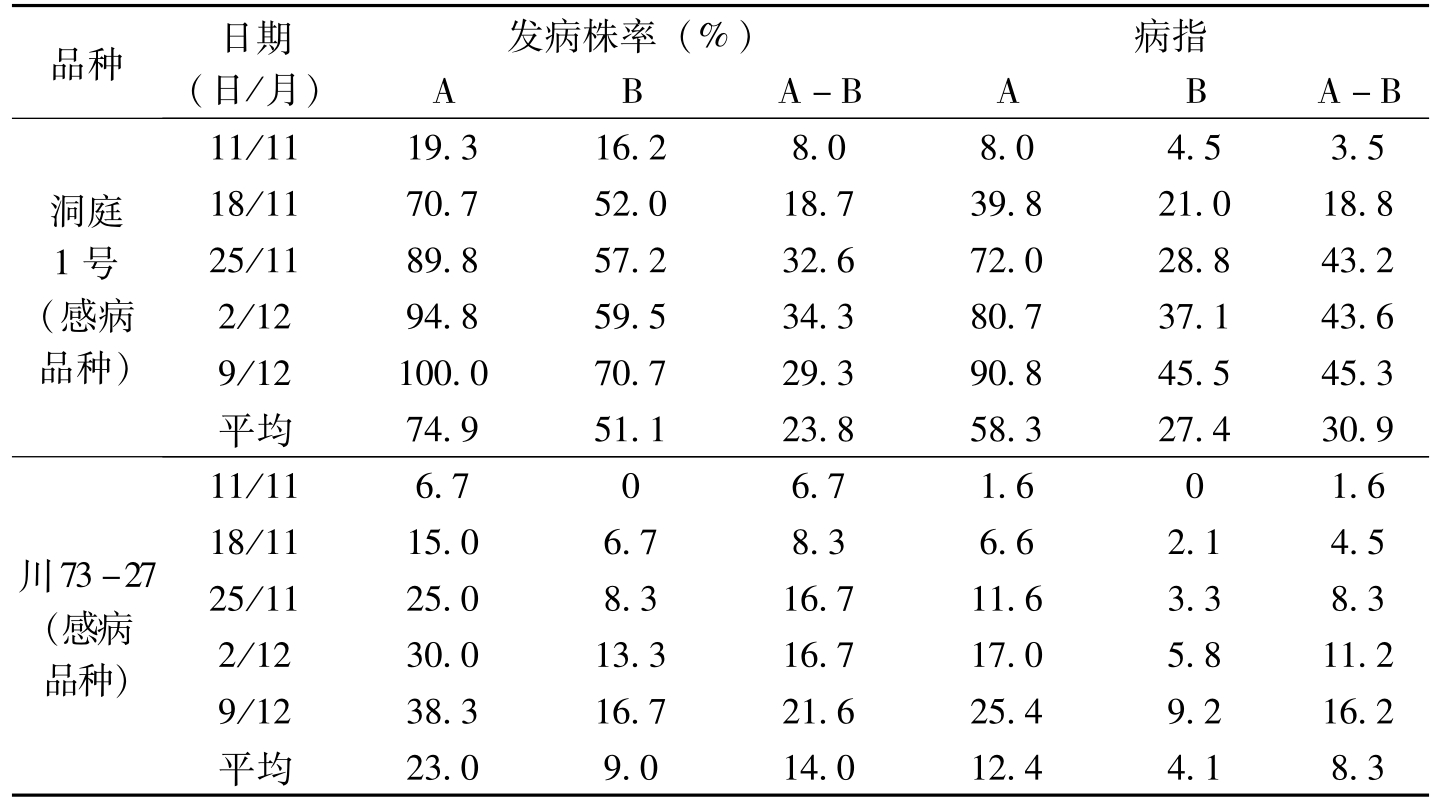
表1-9 棉花枯萎病发病株率和病指
(五)与种子带菌的关系
枯萎病棉株上的棉籽内部带菌问题,已为国内外研究所证实。Elliot(1923)首先肯定棉籽可以携带枯萎病菌。Taubenhans于1929~1931年在分离5094粒棉籽中,带菌率为5.01%。枯萎病随棉籽调引而传播,已被生产实践所证实。追溯我国各地棉花枯萎病最初传入和逐步扩散的历史,不难发现该病大多是由国外引种或从调进外地病区棉种开始的。1935年从美国引进大批斯字棉4B种子,未做消毒处理,就分发到陕西泾阳等处农场和农村种植,这些地方后来就成为我国枯萎病发病最早和最重的病区。在生产上,棉花枯萎病往往是先在引种带病棉种单位的田里发生,然后又随棉种的推广而蔓延为害。
过崇俭等(1957)分离岱字棉14种子时,观察到种胚带枯萎病菌率为1.6%,种子表面带菌率为1.4%;1971年又分离4740粒棉籽,带菌率为0.1%,种子表面带菌率为0.3%,胚叶带菌率为0.6%,胚根带菌率为0.3%。顾本康等(1975)分离抗病品种陕401棉籽15646粒,带枯萎病菌率为0.01%;感病品种岱字棉15带枯萎病菌率为0.1%。可见,无论是抗病品种还是感病品种,只要收获自病田的,均可能带菌,只是带菌率高低不同而已。陈吉棣等于1964~1999年分离11505粒棉种,带菌率为0.026%;仇元等(1963)多次分离棉籽的带菌率为5.9%~39.8%,表明棉籽上可带有多量的病原菌。Evans(1966)分离后认为,未脱绒的棉籽最易粘附病残体,每粒棉种上平均约有10个微菌核。
中国农业科学院棉花研究所(1972)在未种过棉花的黄河故道上进行的试验结果表明,从枯萎病田混收的棉种,其发病率约0.7%;从病株单收的棉种,其发病率为2.2%;而经硫酸脱绒后,其发病率下降97%。这说明种子带菌主要在棉籽的外部,特别是在棉籽的短绒上;但经硫酸脱绒的棉籽,仍有0.23%的棉株发病,间接地证明了棉籽内部可能带有少量的枯萎病菌。
籍秀琴等(1980)进一步研究了棉花种子带镰刀菌种类问题。对8350粒棉花种子进行分离的结果表明,仅镰刀菌即有8种,出现频率较多的有半裸镰刀菌(
研究单位年份种子数(粒)带菌率(%)种子来源中国农业科学院植物保护研究所19647200.14辽宁省辽阳江苏省农业科学院197147400.101江苏省南通中国农业科学院植物保护研究所197531600.15~2.68河南省新乡中国农业科学院植物保护研究所19781506.6陕西省泾阳
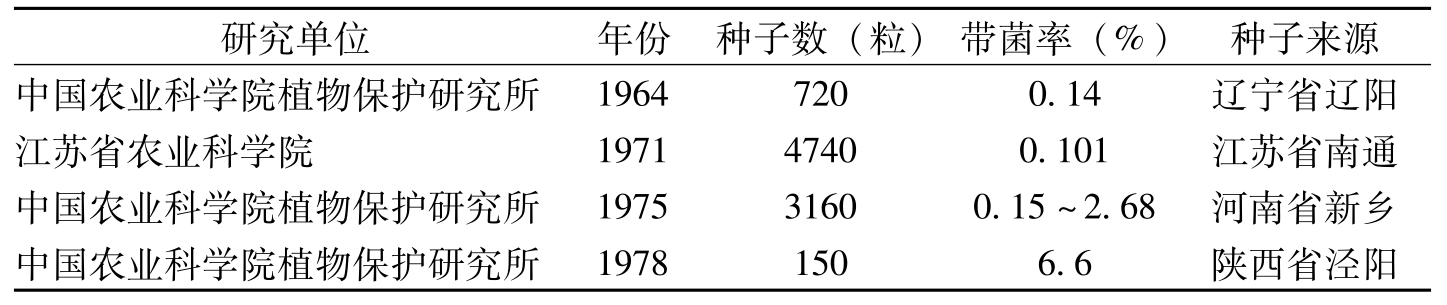
表1-10 不同单位研究的棉花棉籽带枯萎病菌情况
自20世纪90年代以来,尽管我国推广经硫酸脱绒后再包种衣剂的包衣棉籽对减轻因种子带菌而发生的枯萎病有较好的效果,但棉籽带菌的客观事实,仍不容忽视,切实按照国家有关法规的规定,搞好棉籽的检验、检疫与调运,势在必行。
(六)与病株残体的关系
棉花枯萎病病株的根、茎、叶等残体均带有枯萎病菌,这些病残体成为第二年的病菌主要来源。早期发病的病叶及残枝,落在田间也可以引起当年的再侵染。
黄仲生等(1979)进行棉花枯萎病株残体传病试验,施用病叶和病秆沤制的堆肥,枯萎病率为84.1%;施用病叶、病秆喂猪所积的粪肥,枯萎
关于棉田内病残体的测定方法,有间接证明和直接土样测定两种,后者是因为有了选择性培养基,可以从土壤内直接分离病原菌。20世纪50年代,过崇俭等在江苏三余棉场,从枯萎病重病田内,按0~10cm,10~20cm、20~40cm、40~60cm的不同土层,取土置于盆钵内,播种消毒的感病棉种,显症后调查发病率,结果依次为17%、19.6%、10.8%和11.9%,表明40~60cm处的病土仍可引起棉株发病。采用病土播种,土壤内按不同土层掩埋病株体后再播种,均可间接证明由于病土内存在着病原菌而导致棉株显症发生病害。顾本康等(1977)用选择性培养基,在病田内按5点取样法,从表土,10cm、20cm、30cm、40cm、50cm和60cm处取样,经5次取样分离后的平均数,每克土样的棉枯萎病菌的菌落数依次为16.7cm、22.2cm、22.2cm、11.1cm、5.6cm、0cm和0个。证明棉花枯萎病菌可在土壤40cm处的较深层存活,也充分显示棉花枯萎病菌一旦入土,就难以采用土壤处理消灭为害。
枯萎病残体内病菌的存活时间,与它所在条件有关。干燥或水浸对病菌存活均不利,深埋土层20cm内的病株残体内的病菌,19个月存活率才有降低的趋势,而在土表或表土6~7cm内的病菌经18个月就全部死亡。在室内干燥的条件下,13个月不再存活;倘若浸泡水中,则病菌易于死亡,如在室温条件下,病株组织浸于水中46天,病菌存活率较对照降低68%,浸于泥水中降低到80%~90%;如果置于25℃定温条件下,在水中存活率降低到60%~90%,而在泥水中则全部死亡。综合防治中提倡稻棉轮作,或病田泡水其道理即在于此。
五、棉花枯萎病发生的遥感监测
遥感技术的发展和应用为作物的长势监测、种植面积调查和产量估算提供了一个新的科学手段。高光谱遥感技术以其较高的光谱分辨率在植物生物物理参数的遥感定量研究和产量估计中已表现出巨大的应用潜力。利用高光谱遥感技术可以快速精确地获取作物生长状态和环境胁迫的各种信息,是未来精准农业和农业可持续发展的重要手段(陈永芳等,2002)。
中国在棉花高光谱遥感上的研究,多集中在棉花光谱反射率(或导数光谱反射率)与棉花的农学参数,生理参数和水、氮胁迫的相关性分析。唐延林等(2003)研究发现,随着发育期的推移,棉花冠层光谱反射率在可见光范围内降低,在近红外区域增高;叶面积指数、鲜叶重、干叶重和叶绿素含量等与红边的位置、幅值、面积均存在显著的相关性。王秀珍等(2004)指出应用导数光谱法研究棉花光谱特征可以有效地消除土壤背景的影响,在寻找特征波长方面具有广阔的应用潜力。黄春燕等(2003,2006)研究发现,748 nm波段处的一阶微分光谱值与棉花干物质积累量有较高的相关性;反射光谱“红谷”区的数值积分面积和最低反射率与叶片光合速率呈显著的线性负相关,并指出用“绿峰”的偏移作为棉花功能衰退的指标是不可取的。柏军华等(2006)以棉花全生育期叶面积指数与棉花产量的关系和近地高光谱遥感参数模型监测的多时相叶面积指数,可以很好地预测棉花产量。王登伟等(2003)指出棉花一阶微分光谱值与棉叶叶绿素浓度有很高的正相关性,群体叶面积指数与ND-VI呈很好的对数相关性,红边的积分面积与冠层叶片的全氮含量有很高的正相关性。冯先伟等(2004)发现,可见光和近红外波段光谱反射率能够反映出棉花生长发育的动态特征,棉花的花铃期是高光谱遥感对棉花长势和生理参数定量诊断的最佳时期。
棉花枯萎病可以造成棉叶皱缩、失水、叶片紫红色或出现黄色网纹,使棉花的生理参数发生较大改变,这种变化必然会反映在棉叶的光谱曲线上。因此,乔红波等(2007)在温室条件下,利用便携式高光谱仪研究不









