棉花枯萎病病原菌
棉花枯萎病病原菌
一、病原菌的分类学鉴定
导致棉花枯萎病的病原菌为尖孢镰刀菌萎蔫专化型 [
中国一些研究者在分离棉花枯萎镰刀菌研究中,注意到有不同类型大分生孢子,除典型的
二、病原菌的形态特征
棉花枯萎病菌为尖孢镰刀菌萎蔫专化型,其培养性状,在PDA(马铃薯葡萄糖琼脂)培养基上,菌丝为白色,若培养时间稍长培养基经常出现紫色,菌丝体透明,有分离。具有3种类型孢子,分别为大型分生孢子、小孢子和厚垣孢子(图2-1)。镜检时,重点检查有无镰刀状大型分生孢子和厚膜孢子。大型分生孢子呈镰刀形,略弯曲,两头稍尖,足孢明显,无色,大多具3个分隔,少数为2个,偶见有4~5个分隔,大小为(22~23)μm×(3~5)μm(以3个隔膜的大型分生孢子长度和宽度作标准)。小分生孢子呈卵圆形或纺锤形,无色透明,大多数为单胞,少数有一个分隔,大小为(5~6)μm×(2~3.5)μm。厚膜孢子近圆形,顶生或间生1~3个,但也有数个相连的,卵黄色,直径5~15μm,厚膜孢子是抵抗不良环境的繁殖体。
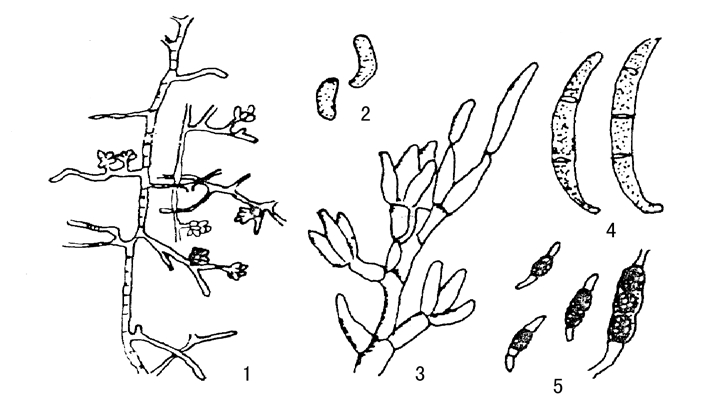
图2-1 棉花枯萎病菌
三、病原菌的寄主植物
自Alkinson 1892年首先提出引起棉花枯萎病菌是侵染导管萎蔫镰刀菌(
许如琛等(1964)用人工接菌盆栽试验,甘薯、玉米、薄荷、黄麻、烟草、荞麦、小麦、大麦、黑麦、燕麦、大豆、豌豆、蚕豆、四季豆、胡萝卜、黄瓜、萝卜、莴苣、油菜、菠菜、青菜和婆婆纳等共47种植物因均不发病,故没有作为枯萎病菌的寄主植物。
顾本康等(1979)将生长在枯萎病菌人工接菌土内的供测作物植株、采用组织捣碎稀释分离的方法,能够分离出枯萎病病原菌,将它回接到棉花上能够表现枯萎症状,再从菌落形态及孢子大小,都证明是棉花枯萎病病原菌,因此认为,玉米、牛角椒、大麦、元麦、小麦、番茄、乌豇豆、茄子、大豆、柽麻、芝麻、花生、山芋、赤豆和扁豆等作物虽然没有明显症状,而确是棉花枯萎病菌的带菌植物。棉花枯萎病菌在很多植物上只能停留于植物的根表皮,但不能深入导管系统,所以,往往出现隐症状态。因此认为,除已被证明的许多作物或杂草是棉花枯萎病菌的寄主植物外,实际上还有更多植物是棉花枯萎病菌的不表现症状的带菌体;这就是在枯萎病区推行与旱作物轮作对防治枯萎病无效的道理。
李君彦等(1990)对小麦、大麦、玉米、高粱、甘薯、大豆、豌豆、红麻、向日葵、烟草、番茄、茄子、辣椒、黄瓜和笋瓜15种植物作棉花枯
四、病原菌致病力分化的鉴定
植物病原菌在不同环境、营养条件和寄主植物影响等外界因素作用下,再加上自身的遗传变异,致病力不断地变化。但在长期演变过程中逐渐形成了一些比较稳定的类型,这就是在种和变种以下所区分的转化型、生理型或小种。专化型是一个群体,据不同鉴别寄主的致病反应,可以进一步化分为若干生理小种。
鉴定棉花枯萎病生理小种,以及生理小种在不同地区的分布,对培育棉花抗病品种,以及优良品种在不同种植区的分布有指导意义。
(一)鉴别寄主鉴定
棉花枯萎病菌具有较强的专化性,不同地区的枯萎病菌尽管形态相似,但致病力不一定相同。Elbeles(1975)根据枯萎病菌对陆地棉、海岛棉、亚洲棉及某些作物的侵染能力的差异,划分为5个生理小种:小种1号和2号,来源于美国,严重感染陆地棉,轻度感染海岛棉,不感染亚洲棉,但小种2号尚能感染烟草及Yel-redo大豆;小种3号为来自埃及的枯萎病菌,感染海岛棉和亚洲棉,不感染陆地棉;小种4号为印度的枯萎病菌,只感染亚洲棉,不感染海岛棉和陆地棉;小种5号为苏丹的枯萎病菌,感染亚洲棉,不感染海岛棉及陆地棉。Armstrong等(1978)描述并提出了巴西的棉花枯萎病菌小种6号。至此,全世界报道的棉花枯萎病生理小种共6个。
1972~1973年全国棉花枯萎病、黄萎病综合防治研究协作组收集了我国主要枯萎病区(冀、豫、晋、陕、鲁、皖、苏、浙、鄂、川等省的县)有代表性的菌株76个,在海岛棉、陆地棉和中棉3个棉种的9个鉴别寄主棉花品种上测定其致病力。结果菌株被划分为3个致病型(表2-1):Ⅰ
鉴别品种致病型Ⅰ致病型Ⅱ致病型Ⅲ海岛棉SSS陆地棉SSR中棉SRR

表2-1 全国各地棉花枯萎病菌在鉴别寄主上的接菌反应
陈其煐等(1985)于1982年用国际通用的一套鉴别寄主对中国各地采集的144菌系筛选出有代表性的17个菌系进行全面的研究,发现我国的棉花枯萎病菌致病力与当时国际上已报道的6个小种有区别。为此,将我国的棉花枯萎病菌分为3个小种,即3号、7号和8号,其中7号、8号小种是首次报道,并认定棉花枯萎病Ⅲ型即为国际3号小种,而Ⅰ型、Ⅱ型被定为新小种,即7号和8号。7号小种是我国的优势小种,广泛分布于国内的各主产棉区,对鉴别寄主中的海岛棉、陆地棉和亚洲棉均表现出高度侵染,不感染或轻度感染5个非棉属寄主;而8号小种则不感染或轻度感染3个棉种的7个品种,轻度感染非棉属的秋葵、金元烟和大豆,严重感染紫苜蓿和白肋烟,仅在我国湖北省的新洲县和麻城县及江苏省的南京发现;3号小种严重感染海岛棉的Coastland、Sakel和亚洲棉的Ronzi,不感染海岛棉的Ashmouni和陆地棉,不感染非棉属寄主的秋葵、金元烟、白肋烟和大豆,极轻度感染紫苜蓿(表2-2)。
鉴别寄主小种及分布3新疆麦盖提和吐鲁番7全国各地8湖北麻城及江苏南京海岛棉AshmouniRSW-RCoastlandSSWSakelSSW
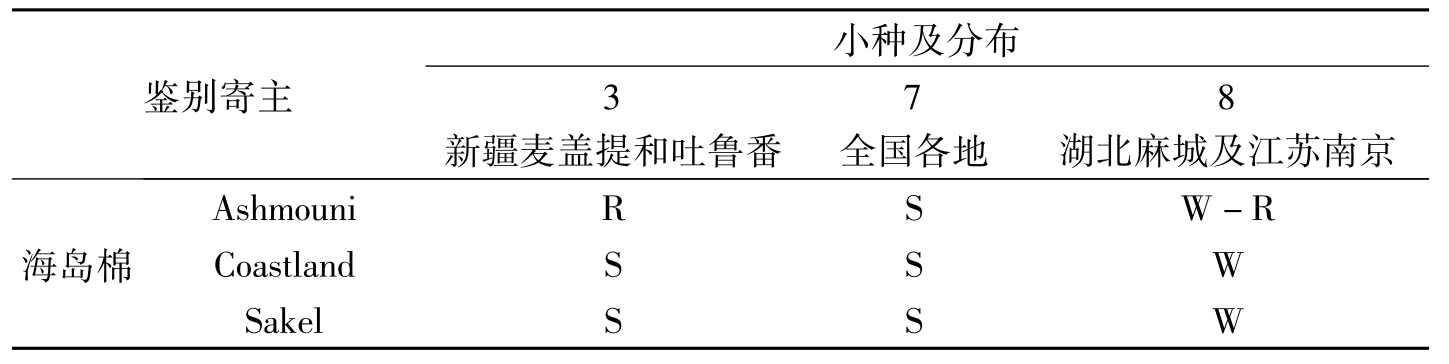
表2-2 我国不同小种对鉴别寄主植物的侵染力(陈其煐等,1985)
鉴别寄主小种及分布3新疆麦盖提和吐鲁番7全国各地8湖北麻城及江苏南京陆地棉AcalaRSWRowdenRSWStonevileRSW亚洲棉RoziSSW秋葵ClemsonspinelaesRW-RW紫苜蓿GrimmRW-RS烟草Burley5RW-RSGolddolarRRW大豆YelredoRW-RW-R
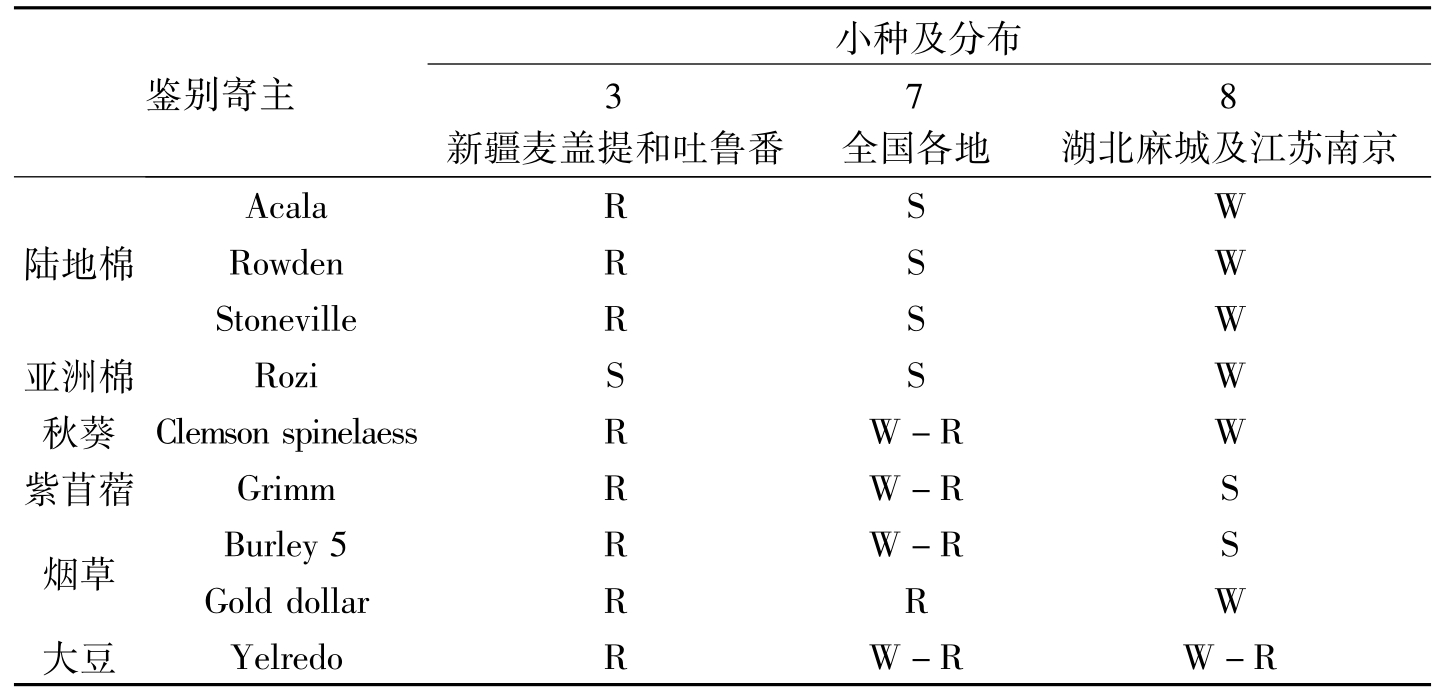
表2-2 我国不同小种对鉴别寄主植物的侵染力(陈其煐等,1985)(续)-1
致病性是病原、寄主和环境条件相互作用的结果。棉枯萎菌致病性变异的原因比较复杂,可能有:镰刀菌发生突变、异核现象和准性生殖而发生变异;棉花种和品种的更换,造成寄主基因型对病原菌群体遗传结构的影响而发生变异;随土壤中微生物群落变化而发生变异及棉枯萎菌靠寄主种子远距离传播,导致外来致病菌的传入或为新小种提供亲本等。为此,孙文姬等(1999)采用国际通用的鉴别寄主(简称通用寄主)包括棉属寄主海岛棉的SaKel、Coastland、Ashmouni;陆地棉的Rowden等4个;非棉属寄主紫苜蓿(Grimm)、白肋烟(Berley5)、金元烟(Gold dollar)3个,以及鉴于国际通用的鉴别寄主中的所有棉属寄主均被7号小种菌系高度感染,不能明显区分广泛分布于我国各棉区7号小种不同菌系间的致病力强弱,于是在1996~1997年试验中,增加了抗病性不同的辅助棉属鉴别寄主(简称辅助寄主),包括海岛棉的8763依(感病)、K102(抗性敏感);陆地棉的鄂荆1号(感病)、86-1号(抗病)等4个品种,对1986~1997年在我国河北、河南、山东、山西、安徽、湖北、江苏、陕西、辽宁和新疆10个主要产棉省(自治区)采集的84个棉枯萎镰刀菌代表菌系进行了生理小种变异监测研究。结果表明:
(1)总体上生理小种类型和分布
与陈其煐等(1985)报道的基本相同,仍为第3号、7号、8号3个
(2)72个7号小种菌系
分布于黄河、长江流域及新疆广大棉区,对非棉属寄主表现为“R”(发病株率为0)或“W”型(发病株率为50%以下),在棉属寄主上以“S”型(病指为20.1以上)为主。1996~1997年28个代表菌系在8个棉属寄主上测定结果,除2个菌系对Sakel致病力为“W”型(病指为20.0以下)外,其余26个菌系对Ashmoumi、SaKel、Coastland、Rowden、8763依、鄂荆1号等6个棉属寄主致病力均为“S”型;28个菌系在有一定抗性的K102寄主上,13个为“W”型,15个表现为“S”型;在抗病品种86-1寄主上,24个为“W”型,4个为“S”型。按各菌系在8个棉属鉴别寄主上平均病指划分为强、中、弱3个类群,4个菌系为强致病类型,17个菌系为中等致病类型,7个菌系为弱致病类型。1998年发现湖北新洲的1个菌系和山西太原菌系对抗病品种86-1的致病力接近“S”型。
(3)1983年分离出3号小种2个菌系
1988年分离的3号小种2个菌系和1991~1994年分离的3号小种变异型5个菌系均分布于3号小种的原发生地新疆吐鲁番地区。对非棉属寄主均为“R”或“W”型;对陆地棉Rowden均为“R”型(病指为0),对海岛棉Coastland均为“S”型。2个3号小种与5个3号小种变异型的差异表现在对Ashmouni和SaKel的致病力上,前者在Ashmouni和Sakel上分别为“R”和“S”型;后者对Ashmoum的致病力增强了,除l个为“R”型外,2个为“W”型,2个为“S”型,而对SaKel的致病力减弱了,均为“W”型。另外,1995~1998年从该地区采集的海岛棉病株上一直未分离到3号小种。
(4)1983~1985年鉴定出8号小种5个菌系
1988~1995年分离的8号小种5个菌系均分布于其原发地区的湖北新洲地区,在非棉属鉴别寄主紫苜蓿和白肋烟上均表现为“S”型,4个在棉属鉴别寄主上表现为“R”或“W”型,其中,1995年分离的Ag136菌系
2007年,马存等把目前世界上已报道的棉花枯萎病菌8个小种归纳成如表2-3所示。
寄主植物小种编号12345678分布地区世界各地美国埃及印度苏丹巴西中国中国亚洲棉(G.arboreumcv.Ronzi)RRSSSRSR海岛棉阿西莫尼(G.barbadebsecv.Ashmouni)SSRRSSSR海岛棉萨克耳(G.barbadebsecv.Sakel)SSSRSSSR陆地棉阿卡拉44(G.hirsutumcv.Acala44)SSRRRSSR金元烟(Nicotianatabacumcv.GoldDolar)RSRR—RRR大豆(Glycinemaxcv.Yelredo)RS——RRR羽扁豆苜蓿(Lupinusluteuscv.Weiko)SS——R
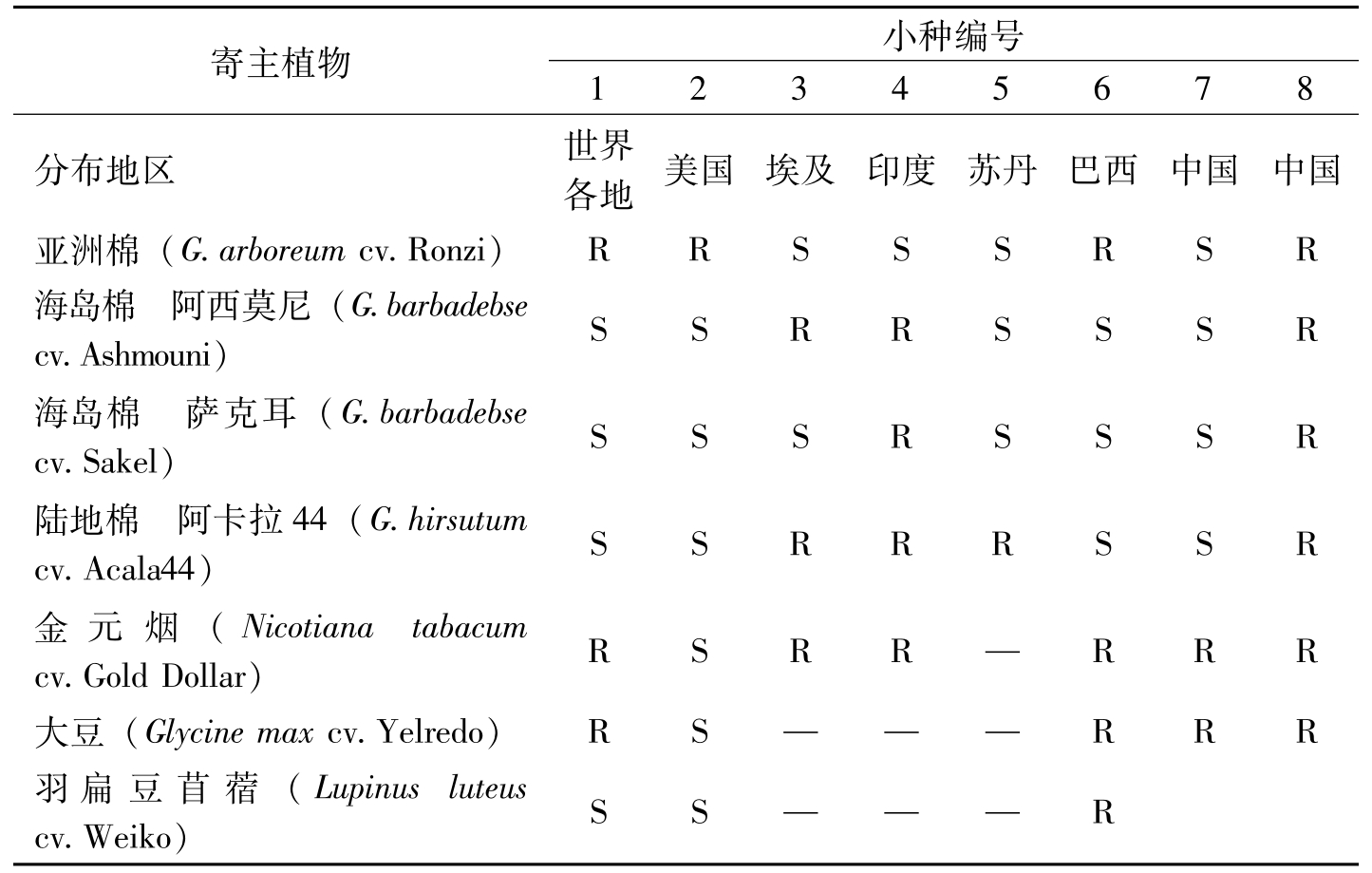
表2-3 世界各国不同小种对不同寄主植物的侵染力
新疆棉区棉花枯萎病生理小种,自徐怡心(1994)鉴定认为国际7号小种之后,李国英等(1998)、缪卫国等(2000)、王雪薇等(2001)、吴彩兰等(2004)和张莉等(2005)采用目前国际上通用的一套棉属鉴别寄主及当地的辅助鉴别寄主对从不同产棉县(市、团场)采集的枯萎病菌株样本鉴定后认为,目前,7号生理小种仍为新疆棉花枯萎病菌优势生理小种,主要分布在新疆南疆(和田、喀什、阿克苏)、北疆(石河子、昌吉、乌苏)、东疆(吐鲁番)主要棉区。从各供试菌系对9个品种的发病始期及发病程度看,新疆棉花枯萎病菌菌系强致病型主要分布在南、北疆主要棉区,部分菌系属中等致病型,枯萎病菌为害时期较早,棉株出苗后10~12天,造成棉苗子叶青枯萎蔫或叶脉黄化网纹;新疆棉花枯萎病菌菌系弱致病型主要分布在东疆棉区,枯萎病菌为害时期均较晚,较南、北疆棉区
(二)营养体亲和性鉴定
营养体亲和性(Vegetative Compatibility,简称VC)是指缺乏有性生殖真菌,通过不同菌系间的菌丝融合交流物质,进而形成异核体的能力,其实质就是异核体亲和性,会导致病原菌形态和致病性的变异。由若干具有营养体亲和性的菌系构成的群,称为营养体亲和群(Vegetative Compatibility Group,简称VCG)。
做致病性测定存在着费时、费力、结果易受外界因素影响及不能反映不同菌株间的遗传关系等公认的问题(Hart等,1981;Kraft等,1978;Pound等,1953),所以,有必要探索和寻找鉴别小种更为简便有效的途径。Puhalla(19850)首先确立的用不能还原硝酸盐作唯一氮源生长的突变体(nit突变体)作营养体亲和性试验的方法,为此提供了一条有效的途径。此法通过观察不同菌株的nit突变体之间能否互补而形成异核体把它们分成不同的营养体亲和群。根据他的理论,亲和与否反映出不同菌株在遗传进化过程中的亲缘性远近,国内外不少报道都认为营养体亲和性是鉴别镰刀菌不同专化型及小种间菌株的有效方法。
1985年Puhalla首次将VCG技术应用到尖孢镰刀菌专化型的研究,结果认为专化型与VCG之间有一定的对应关系,即同一VCG的菌系属于同一专化型,而不同专化型的菌系属于不同VCG。随后,大量的不同专化型的菌系证明了Puhalla的观点(Ploetz,1990)。鲍建荣等(1992)对尖孢镰刀菌的6个专化型的20个菌系进行了营养体亲和性研究,结果显示,不同专化型菌系的nit株之间无互补反应,而同一专化型的不同菌系上分离的nit株之间可产生亲和反应,也证明了nit突变株的营养体亲和群与菌系的专化型有相关性。在棉花专化型内,研究证明,生理小种与VCG有一定的相关性,来自同一生理小种的菌系,甚至是不同地理来源的菌系,都可划为同一VCG。Katan等(1988)测定来自以色列全国不同地区的同为3号生理小种的12株棉花枯萎病菌菌系,结果均属于同一VCG。丁之金等(1992)采用不能还原利用硝酸盐作唯一氮源生长的突变体(nit突变体),
营养体亲和群(VCG)菌株数菌株来源营养体亲和群(VCG)菌株数菌株来源VCG15江苏丹徒VCG26江苏丹徒(303,315,316,319,328,329)江苏江浦江苏启东江苏淮海农场VCG31江苏丹徒(234)新疆四川浙江VCG41江苏南京(402)江西
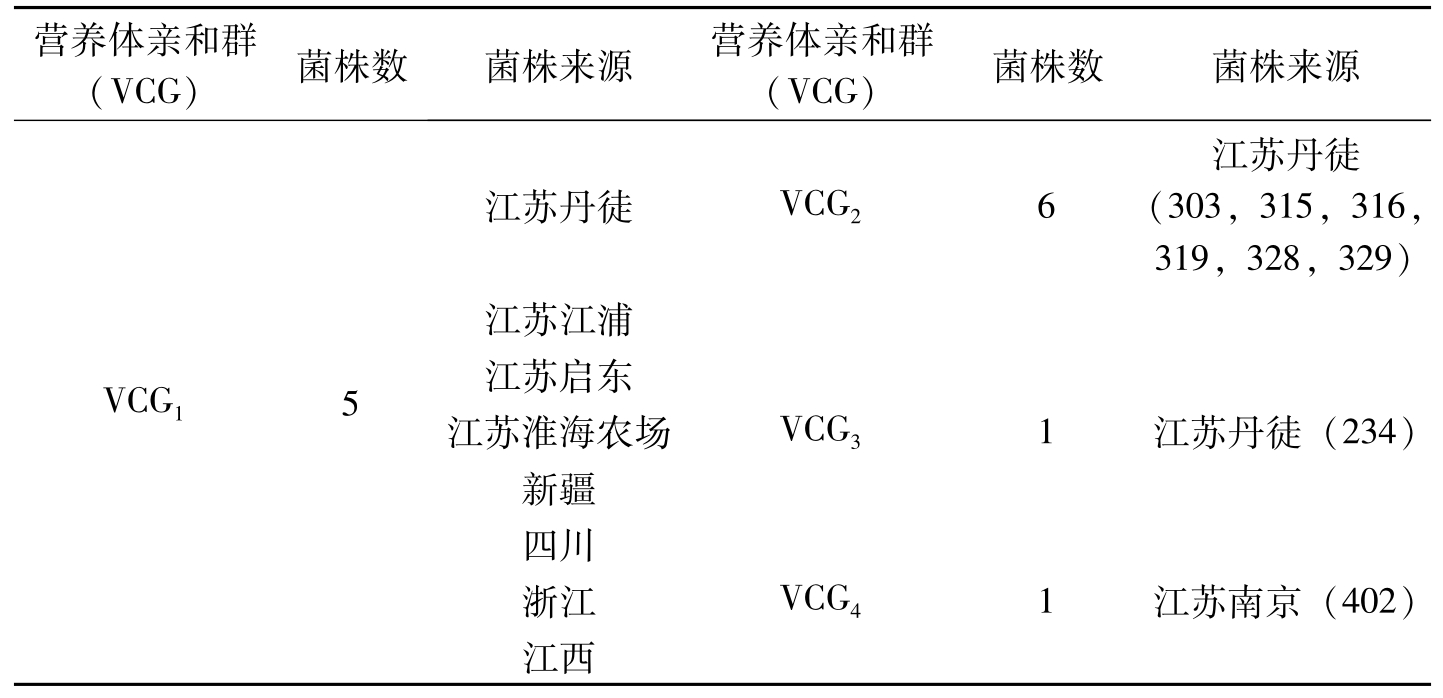
表2-4 棉花枯萎病菌的营养体亲和群及其来源
除了属于4个VCGs的103个菌株,另有4个菌株(菌株号219,226,227,302)只产生一种表观型的突变体(nit A),这些突变体与其他菌株的互补型突变体配对不能形成亲和性反应。这些实验结果表明,我国的部分地区,包括新疆、四川、浙江、江西和江苏的棉花枯萎病菌多属于同一个营养体亲和群。在表现症状的棉花上部茎叶中也分离到其他营养体亲和群的菌株,这些菌株经测定,对陆地棉的棉花品种具有寄生能力和一定的致病力。
王雪薇等(1996)报道,新疆棉花枯萎病株或病田土中分离得到的尖孢镰刀菌具有明显的营养体亲和性。将分离自新疆5个不同地点的棉病株或棉田土的7个菌株经单孢分离得到18个单孢株,经鉴定均为尖孢镰孢。7个菌株的18个单孢株分别在KPS培养基上诱得数量不等的nit突变体,将它们分别与两个采自老病区莎车的棉枯萎病菌株进行营养体配对试验,结果显示,7个菌株分别归属2个营养体亲和群(VCGs),其中,与莎车菌株明显亲和,属V1的6个菌株与莎车菌株应同属于尖孢镰孢萎蔫专化型。研究结果还表明,不同菌株产生的亲和带的形态特征不同,如MK菌株的各nit突变株间产生的亲和带均表现为水平扩展,迅速呈纺锤形,但气生菌丝不发达,亲和带仅表现为菌丝层比突变体菌丝层厚,随着亲和带的发展可在两突变株间连成片,暂称扩展型亲和带;而MK7-1菌株的各nit突变株间产生气生菌丝茂密,且扩展宽度均匀的线形亲和带,亲和带朝更浓密方向发展,仅加宽,但不易成片,暂称线型亲和带。所有亲和菌株(或单孢株)与MK形成的亲和带均属扩展型亲和带,而与MK7-1形成的均属线型亲和带。
依据不能利用硝酸盐的突变体(nit)间营养体亲和性划分出的棉花枯萎病菌营养体亲和群(VCGs)与其生理小种间高度相关,即不同生理小种间营养体不亲和,而同一生理小种归属于一个或少数几个VCGs。张莉等(1998)从新疆各植棉区采集棉花枯萎病病株,经分离纯化获49个单孢菌株。经营养体亲和性测定结果表明,供测49个菌株分为4个亲和群,其中,VCG1包括46个菌株,占供测菌株的93.9%,均与标准菌株中的7号小种Ag84亲和,另外3个(吐-2、昌-4、石-2)分属不同的亲和
20世纪90年代后期以来,随着新疆植棉面积的不断扩大及从各地大量调运棉种,其病原种群是否有所变化,不少植棉单位存在一定疑问。为此,张莉等(2005)采用营养体亲和性技术,对新疆棉花枯萎病菌的生理小种类型及其病原种群进行了变异监测研究。结果表明,28个供试菌系高度侵染海岛棉、陆地棉及K102(辅助鉴别寄主),属典型的7号生理小种,其余12个菌系在鉴别寄主上的反应与7号生理小种略有差异;供试菌系属于一个营养亲合群,且与7号生理小种的标准菌系相亲和,与3号、8号小种的标准菌系不相亲和;新疆棉花枯萎病菌依旧以7号生理小种为主。与以往的研究结果相比,新疆棉花枯萎病菌的群体组成基本没有发生变化。此外,王雪薇等(2000)的研究结果也表明,52个待测菌株与7号小种的4个标准菌株间存在着不同程度的营养体亲和性,而与3号小种和8号小种的标准菌株均不亲和。因此,它们属于7号小种营养体亲和群—VCG701。
在棉花枯萎病菌的生理小种及其遗传进化关系的研究中,通过突变体的诱发而进行的营养体亲和群判别方法是行之有效的重要研究方法之一,但在突变体的诱发与鉴定上存在较大的难度,方法不一,试验的稳定性较差。为此,白剑宇等(2007)对棉花枯萎病菌突变体的诱发与鉴定做进一步的方法验证与技术探讨。从32个供试棉花枯萎病菌菌株中共诱得288个nit突变体,根据其在不同氮源培养基上的生长划分出4种突变体类型:nit 1、nit 3、nit M和nit 8。对各菌株的突变体诱发与鉴定结果表明诱得突变体的难易程度主要因菌株的不同有很大差异,有些菌株(FKLMYN-23,FKTN-11,FSYN-Z6)很容易诱得各类型突变体,有些菌株不易诱得某种突变体类型,有个别菌株(FMYN-02,FYL2N-07)在KClO
这些结果对建立稳定和准确的实验方法体系,为进一步查明棉花枯萎病菌的营养体亲和群分化情况,营养体亲和群与生理小种的对应关系,生理小种的种类及其分布,病原菌的分化,遗传进化关系提供了有科学价值的依据。
异核现象是半知菌普遍具有的特性,也是病原菌变异的主要原因。史大刚等(1991)结合新疆吐鲁番地区,原只发现枯萎菌生理Ⅲ型,后又发现了生理Ⅱ型的现象,研究了异核现象与枯萎菌变异的关系认为,异核现象是导致生理Ⅲ型菌系致病力增强产生生理Ⅱ型的重要因素。
棉花枯萎病菌在遗传上都是十分复杂的种群,遗传的多变性可以由多种因子引起,例如,转座因子、染色体突变、基因漂移、准性生殖等。这种种群上遗传多变性决定了必须依靠更为客观的研究方法来研究它。VCG是依靠真菌自身遗传特征来划分的,不仅能反映菌系之间的遗传相似性,而且有助于真菌繁殖方式及其群体遗传结构的分析。但目前,棉花枯萎病菌营养体亲和性研究菌系来源十分有限,而且棉花枯萎病菌主要集中在专化型内,这样不能更为确切地了解种群的发育与变异。因此,今后应进一步扩大棉花枯萎病菌的研究种群,并结合更多的研究技术,包括分子技术,对日益复杂化的棉花枯萎病菌的种群进行更深入的研究。
(三)同工酶技术鉴定
生物体内的同工酶是基因与生物外部形态性状的连接物,同工酶酶谱是生物内部生物化学特性的反映,许多学者认为,病菌不同的致病类型与酯酶同工酶有较密切的关系。吕金殿等(1982)用聚丙烯酰胺凝胶电泳对42个采自我国各省(自治区)的棉花枯萎病菌的酯酶同工酶进行了比较研
(1)新疆棉花枯萎病菌有4种酶谱类型,供试的30个菌株中有25个菌株的酶谱属于类型Ⅰ
据以往研究,这25个菌株中,有24个菌株属于7号生理小种,它们与标准的7号生理小种的酯酶同工酶酶谱基本一致。这说明,可利用酯酶同工酶作为研究棉花枯萎病菌致病力分化的一种辅助方法。同时,从生化的方面再次证明7号生理小种是新疆棉花枯萎病菌的优势小种。
(2)主酶带的多少与致病力的强弱成正相关
类型Ⅰ中的菌株有5条主酶带,次酶带也最多,在供试菌株中属致病力最强的类型;类型Ⅱ中的菌株有2条主酶带,次酶带较少,属供试菌株中致病力较弱的类型。
(3)试验证明,酶谱类型与生理小种的类型并不完全一致
如塔2菌株的酶谱类型属于类型Ⅳ,它与本次试验提供的3个标准菌株酶谱均不相似,但在利用鉴别寄主的鉴定中属7号小种;还有石-15菌株酶谱类型与标准3号小种相一致,但其小种也属于7号。这些现象有待进一步研究。
曹君等(2005)通过聚丙烯酰胺凝胶电泳测定了各供试枯萎病菌酯酶同工酶,并结合同工酶谱的聚类分析研究其酶谱类型与菌株致病力及生理分化的关系。结果表明,在棉花枯萎病菌中酯酶含量较高,电泳酶谱稳定,重复性好。14个供试菌株与标准菌株J-1的枯萎病菌酯酶酶谱基本一致,既反映了棉花枯萎病菌在专化型水平上的遗传一致性,又能在专化型下区分出不同的致病类型。根据酯酶酶谱和聚类分析结果(图2-2)将供试菌株分为4种类型。类型Ⅰ的菌株有5条主酶带,次酶带也最多,在供试菌株中属致病力强的类型;类型Ⅱ中的菌株的致病力属于中等水平,
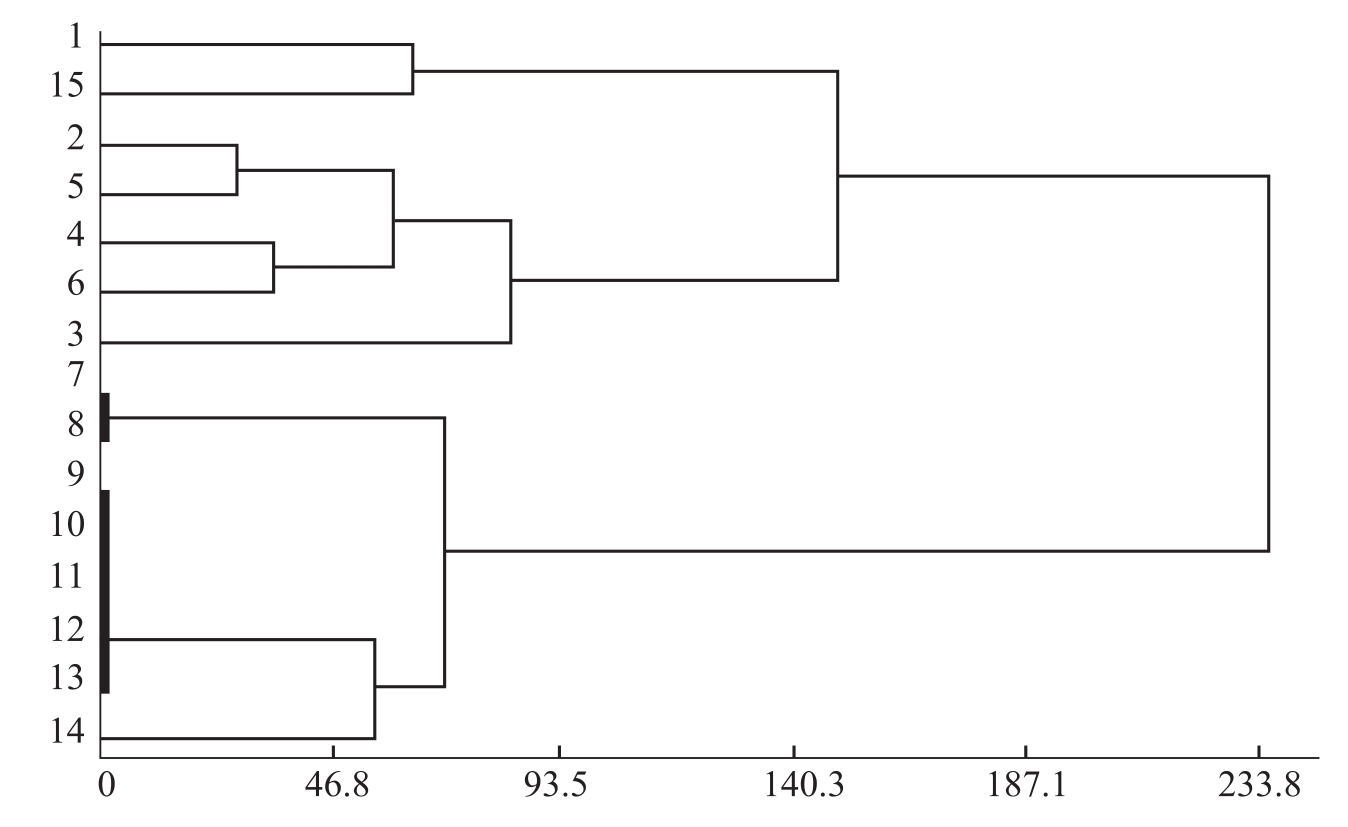
图2-2 同工酶酶谱聚类分析树状图
棉花枯萎病菌各生理小种间,不论是酯酶同工酶、过氧化物同工酶,还是可溶性蛋白质,经电泳后或多或少都可出现特征性条带,差异比较明显,易于区分。吴彩兰等(2004)对棉花枯萎病菌的3号、8号和7号3个生理小种的标准菌株和新疆所采31个供试菌株采用聚丙烯酰胺凝胶电泳进行了酯酶同工酶谱、过氧化物同工酶谱和可溶性蛋白质谱分析。结果表明:①菌体酯酶同工酶电泳:所有供试菌株的酯酶同工酶酶谱多态性丰富,谱带数目一般为3~8条,清晰可辨。棉花枯萎病菌不同生理小种的酯酶同工酶谱存在明显差异,3号生理小种有6条谱带,8号生理小种有3条谱带,7号生理小种有8条谱带,其中,Rf值为0.1和0.35的两条带为7号生理小种的特征性条带。供试菌株谱带数为4~8条,其谱带虽具有多型
吴彩兰等(2004)采用SDS-聚丙烯酰胺凝胶电泳法,对2002~2003年,从新疆各主要植棉区采集到的31个棉花枯萎病菌菌株及3号、7号和8号生理小种的标准菌株进行了可溶性蛋白质谱分析。结果表明,棉花枯萎病菌不同生理小种的谱带存在明显差异,易于区分,并且3号和7号标准菌株都有自己的特征性谱带;同一生理小种不同菌株谱带的一致性较高,差异不大,供试菌株与7号生理小种的特征性谱带中的1条或2条一致。从电泳图谱聚类分析可将其分为3类:3号和8号标准菌株各为1类,供测大部分菌株的电泳图谱与7号生理小种标准菌株的相似性水平高,亲缘关系近,与3号生理小种的相似性水平低,亲缘关系最远(图2-3)。
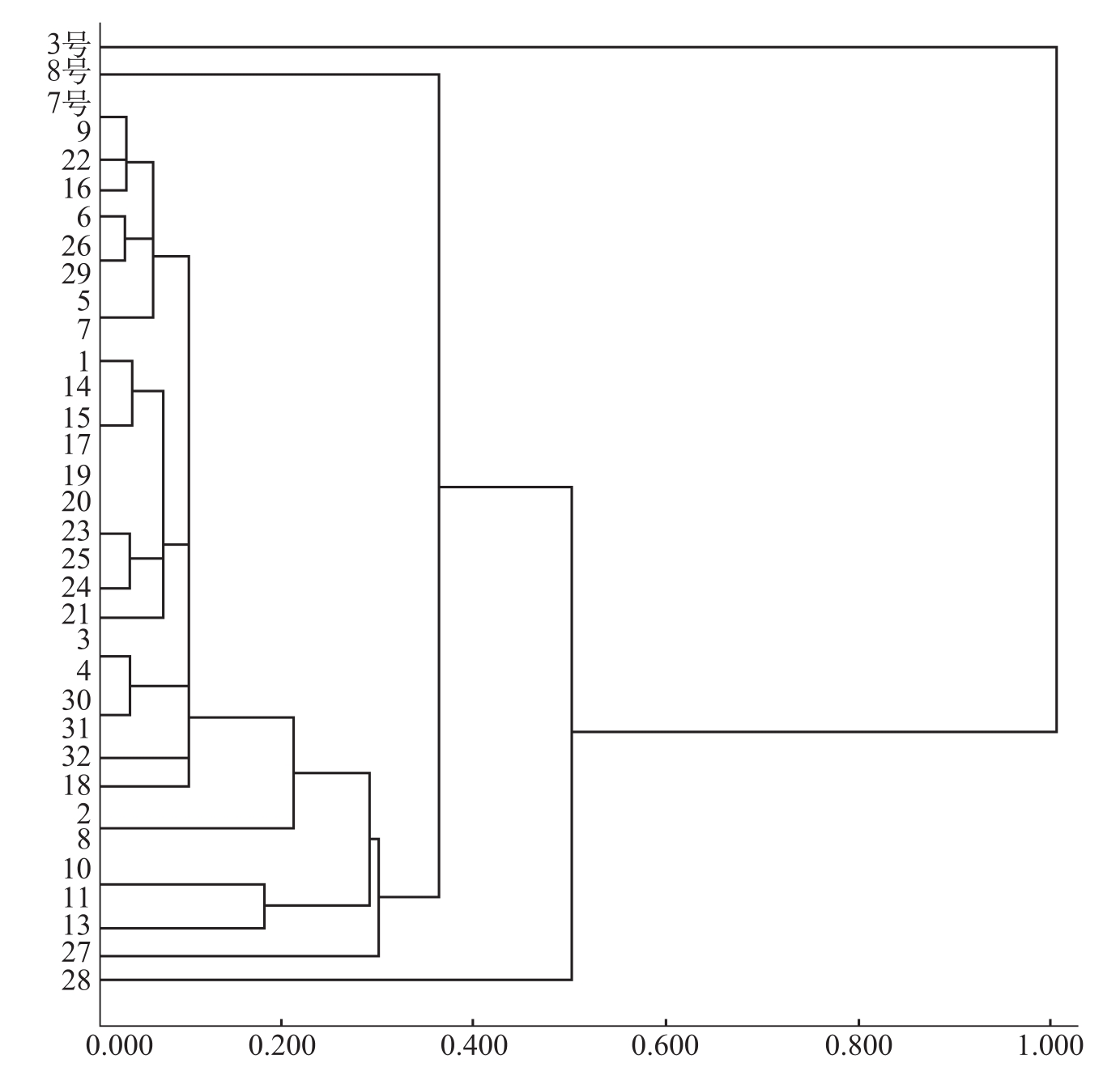
图2-3 棉花枯萎病菌供试菌株电泳图谱聚类分析树状图
(四)分子标记技术鉴定
由于传统的生理小种划分主要以病菌对特定寄主的致病力强弱作为依据,病菌自身的变异及环境因子的复杂变化也可造成致病力变异,仅依靠传统的形态学和致病力差异来划分病菌生理小种已难以得出令人信服的结论。因此,需要借助于诸如遗传学、生物化学、分子生物学的手段加以验证,如血清学、营养体亲和性、凝胶电泳以及近年来发展迅速的RFLP及
RAPD(Random Amplified Podymorphic DNA)技术是20世纪90年代由William等在PCR基础上创立起来的一种利用随机引物扩增DNA的分子标记方法。该技术一出现,就以其快速而便于检测大量样品,所需DNA量少、不需要生物的特殊基因文库作探针以及操作安全、所需费用较低等突出优点,迅速应用于生物学研究的各个领域。冯洁等(1999)对来自中国11个省(自治区)的棉花枯萎病菌不同生理小种菌株的基因组DNA进行随机扩增,筛选出10个扩增多态性好且稳定的随机引物,不同引物的条带数为9~19条,扩增片段的DNA分子量为300~3000bp,10个引物扩增得到的140个DNA条带中,有123个多态性条带,占总条带的87.8%。根据供试菌株RAPD-PCR扩增条带的有无,以1.0记数,统计稳定、清晰出现的条带,采用Statistics统计软件对数据进行类平均法系统聚类分析,建立树状图(图2-4)。以连锁距离为5.0划分时,供试的29个菌株可划分为6个RAPD组。Ⅰ组为所有的3号小种菌株(1~5)及国外3号小种对照(29);Ⅱ组为所有的7号小种共16个菌株(11~26);Ⅲ组为8号小种的3个菌株(6~8);Ⅳ组为8号小种的另外2个菌株(9、10);V、Ⅵ组分别为国外的1号小种(27)及6号小种(28),它们各独自为一类。通过亲缘关系树状图可以看出,我国的3号小种与国外的3号小种对照亲缘关系十分密切,同属于一个RAPD组Ⅰ,这个RAPD组与其他5个组的菌株在亲缘关系上相距较远,独为一类;我国特有的7号小种自成一类;而我国的8号小种遗传背景较为复杂,分属于2个不同的RAPD组,其中,归为第Ⅳ的菌株(9、10)与7号小种的亲缘关系较近,归为第Ⅲ组的菌株(6~8)与7号小种的亲缘关系较远。国外的1号、6号小种(27、28)与我国的7号、8号小种在亲缘关系上相距较远。从亲缘关系树状图中还可以看出,我国的3号、7号小种都分别属于两个独立的RAPD组,8号小种分属于
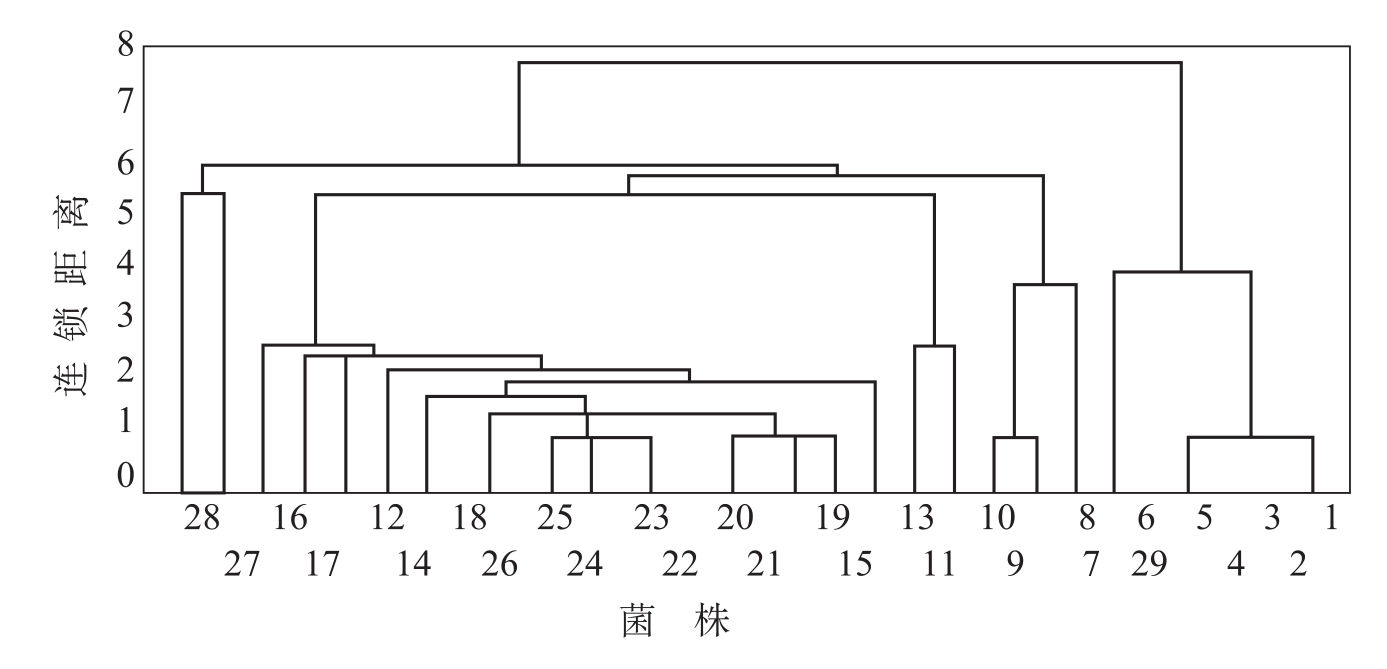
图2-4 供试棉花枯萎病菌菌株的RAPD聚类分析树状图
新疆是我国最大的棉花生产基地,但植棉面积迅速扩大、连作年限延长、引种频繁,导致棉花枯萎病迅速扩展,为害严重。为了有效控制棉花枯萎病菌,有必要对新疆不同植棉区枯萎病菌的生理小种及遗传多样性进行分析,为新疆棉花抗病育种、植物检疫及病害防治提供科学依据。王雪薇等(2001)和田新莉等(2002)利用RAPD技术比较新疆棉花枯萎病菌不同菌株间在分子水平上的共性与差异。RAPD分析结果显示出这37个供试菌株与7号小种各对照菌株间基因组DNA的指纹图谱高度相似,属同一遗传相似组,而与3号和8号小种的对照菌株间遗传差异较大,亲缘关系较远,即7号生理小种是组成目前新疆棉花枯萎病菌群体的优势小种。仅发现一株可能属于3号小种的菌株,未发现属于8号小种的菌株。此外,从分子水平上证明新疆棉花枯萎病菌存在明显的遗传分化,即使在7号小种内也存在遗传上的差异,从而导致致病性强弱的不同。
冯洁等(2000)采用10个随机引物对我国3个棉花枯萎病小种的26个菌株PCR扩增结果表明,不同小种在扩增的DNA条带之间差异明显,
利用随机引物对棉花枯萎病菌不同生理小种的菌株进行PCR扩增,获得了大量的RAPD标记,并分别筛选到可将不同小种扩增出不同特征带的引物,为以致病力差异为基础的鉴别寄主划分生理小种的传统方法提供了分子水平的证据,说明我国棉花枯萎菌不同生理小种间在分子水平上存在较大差异,并且这种差异导致了病菌致病力的不同,不同小种之间特征性条带的存在也正是基因序列改变的真实体现。在棉花枯萎病菌小种间寻找到特异性DNA扩增片段,目前在国内外尚属首例。
AFLP(Amplified Fragmentlength Polymorphism)分子标记技术是在基因序列未知的情况下,利用少数几对引物对相关物种基因组多态性进行有效检测。AFLP、RAPD及RFLP 3种分子标记多态性的检出效率的大小顺序依次为:AFLP>RAPD>RFLP。由于AFLP综合了RAPD和RFLP的技术优点,因而在植物病原真菌的遗传多样性分析、构建遗传图谱、标定基因、辅助育种的研究中得到了广泛应用。王兰等(2008)应用AFLP分子标记技术,选用9对EcorⅠ和MseⅠ引物组合,对新疆南疆地区不同团场的枯萎病菌12个菌系进行AFLP扩增,结果显示,每对引物均扩增出多态
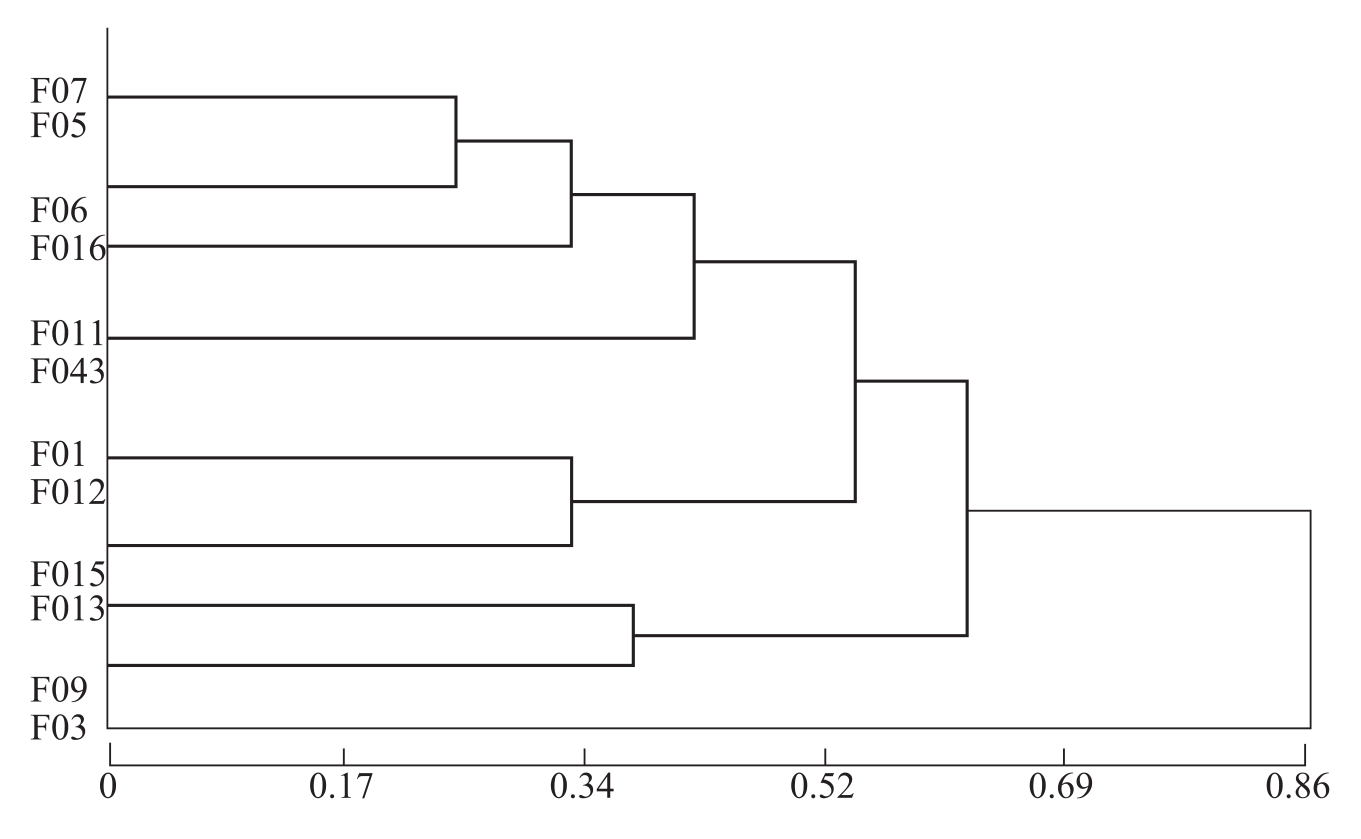
图2-5 12个不同菌株AFLP聚类的树状图
张莉等(2009)研究结果显示,新疆棉花枯萎病菌的AFLP多态性较高,这充分反映了枯萎病菌7号生理小种的群体内存在着一定的遗传多样性。出现这种情况的原因可能有两个:一是棉花枯萎病主要靠种子带菌传播,新疆大量从内地调运棉花种子,外来的棉花枯萎病菌传入新疆,这也暗示着新疆棉花枯萎病菌最初可能不是共同的来源;二是随着抗病品种的应用以及气候条件的变化,当地菌株发生突变,形成了适应新品种和当地自然条件的新菌株。研究的结果还表明,新疆棉花枯萎病菌AFLP多态性
AFLP技术包含多个实验环节,要想获得理想的实验结果,需要对每个实验环节进行优化。在DNA酶切、连接试验过程中,需要制备高质量的DNA:一方面,DNA的纯度影响着酶切的效果;另一方面,DNA的完整性影响着实验结果的稳定性和重复性。单纯固体或液体培养基中含有糖和琼脂等营养物质,培养过程中使棉花枯萎病菌细胞中富含多糖、蛋白质等杂质,不利于DNA的纯化。限制性内切酶的选择对AFLP分析的准确度具有关键性作用,一般采用双酶切,其组合由1个高频内切酶和1个低频内切酶组成。目前国内外大多采用EcoRⅠ/MseⅠ和PstⅠ酶切组合,但Gomez等利用HaeⅢ/PstⅠ酶切组合和EcoR I/Mse I酶切组合对黑莓锈病病菌(
五、影响棉花枯萎病菌生长的因素
棉花枯萎病菌的生长及其产孢量是病原菌生命活动和自身遗传的反应,同时也受到温度、pH值、碳源和氮源等因素的影响。
(一)温度对棉花枯萎病菌生长的影响
李生才等(1998)认为,病菌生长温度范围为10~33℃,最适温度为27~30℃。曹君等(2005)研究结果表明(图2-6),供试棉花枯萎病菌菌株在10~35℃下均能生长,最适温度为25℃。棉花枯萎病菌生长温度因菌株不同而有差异已经得到证实。Raillo(1958)指出,尖孢镰刀菌生长适宜温度15~25℃,最高温度为35℃;陈其煐等(1992)报道,棉花枯萎病菌最适生长温度为25℃,最高温度大多为35℃,但有2个菌株能在37℃下生长;缪卫国等(2000)的研究结果与陈其煐等(1992)报道相似,但同时发现新疆吐鲁番棉枯萎病菌菌株HAI-17较耐高温,能在40℃下生长。不同研究结果所显示的棉花枯萎病菌生长温度的差异可能主要与供试菌株个体差异性有关,后者又与遗传和环境等因子有关。
(二)pH值对棉花枯萎病菌生长的影响
李生才等(1998)报道,棉花枯萎病菌在pH值2.5~7.0均可生长,pH值3.5~5.3为最适。曹君等(2008)研究结果指出(图2-7),棉花
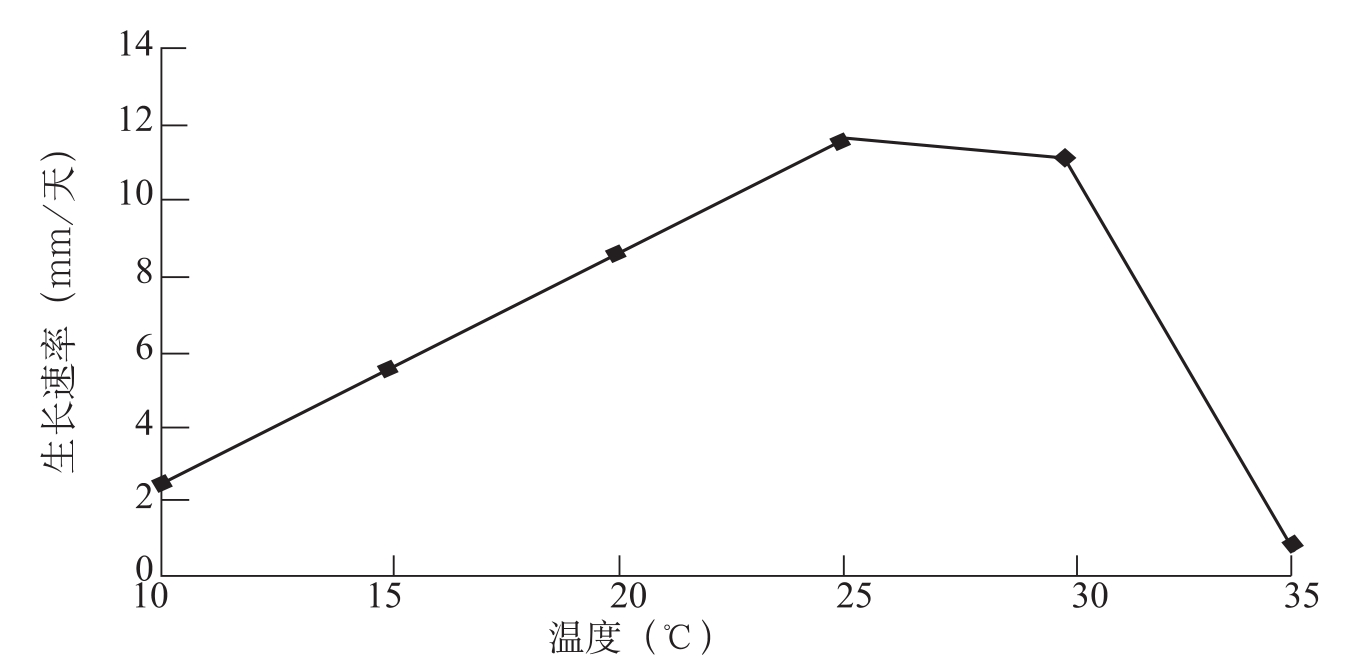
图2-6 不同温度对棉花枯萎病菌菌丝生长的影响

图2-7 不同pH值对棉花枯萎病菌菌丝生长的影响
(三)碳源对棉花枯萎病菌丝生长及产孢量的影响
在供试的4种碳源中,供试菌株在以蔗糖、乳糖、可溶性淀粉为碳源的培养基上菌丝生长较快,3种碳源之间在0.05水平上无明显差异。在以
碳源生长速率(mm/天)菌丝干重(g)产孢量(个/ml)CK(清水)—0.0732.80×106蔗糖12.611.94845.93×107可溶性淀粉12.362.21854.76×107乳糖12.080.29211.20×108甘油11.210.33512.50×107
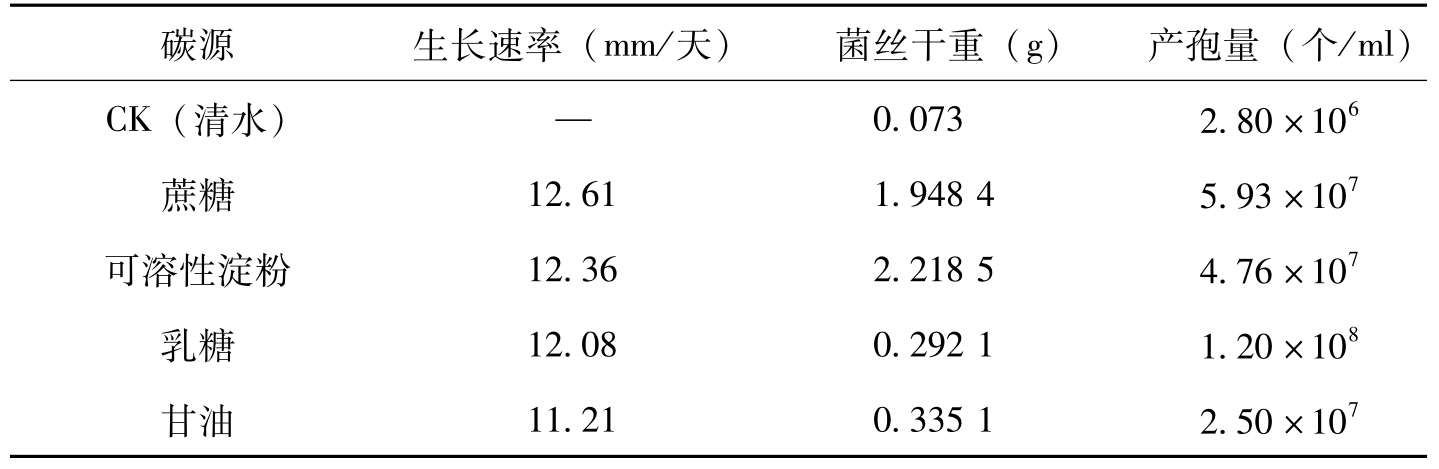
表2-5 碳源对棉花枯萎病菌生长、产孢的影响(曹君等,2008)
(四)氮源对棉花枯萎病菌生长及产孢的影响
在供试的5种氮源中,供试菌株在以脲、硝酸钠为氮源的培养基上菌丝生长较快,两氮源在0.01水平上无明显差异。硝酸铵其次,在0.05水平上与前两种氮源以及磷酸氢二铵、硫酸铵差异明显。磷酸氢二铵、硫酸铵两氮源对菌丝生长影响较小。在供试氮源中,对菌丝干重而言,硝酸铵最好,磷酸氢二铵其次,脲、硝酸钠菌丝干重的差异不明显,硫酸铵较差,无氮处理(CK)的菌丝干重很少,说明氮是该菌生长的必需元素,对该菌的生长必不可少。对于产孢量而言,在供试氮源中以硝酸铵最好,脲、硝酸钠、磷酸氢二铵以及无氮对照其次,硫酸铵的产孢量较少(表2-6)。
碳源生长速率(mm/天)菌丝干重(g)产孢量(个/ml)CK(无氮)—0.06483.21×107脲12.560.20624.28×107硝酸钠12.500.20683.32×107硝酸铵10.670.27301.40×108磷酸氢二铵5.610.22782.24×107硫酸铵3.200.17373.80×106
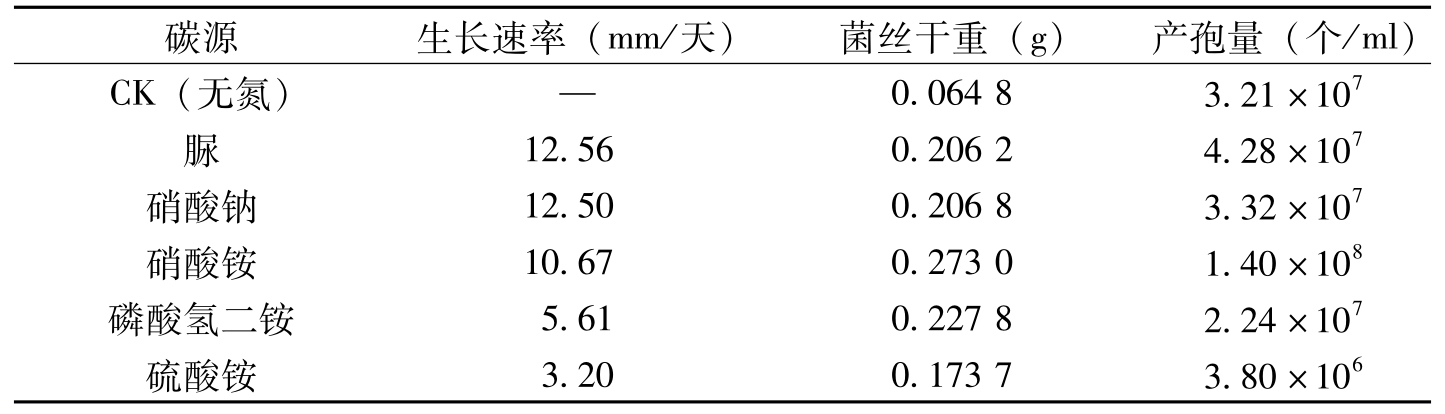
表2-6 氮源对棉花枯萎病菌生长、产孢的影响(曹君等,2008)









