棉花枯萎病的致病机理及抗病机制
棉花枯萎病的致病机理及抗病机制
一、致病机理
对于枯萎病的致病机理,历来有不同的解释。其中,Melhus(1924)、Waggoner等(1954)提出堵塞说,认为棉株感病后导管被堵塞,水分不能向上移动之故。而Fahmy(1923)、Schaffnit等(1932)和Elpidina等(1935)认为,病菌对棉株产生有毒物质,并沿植株上行液流,流至各部,使原生膜透性、碳氮代谢、呼吸作用受损,各种酶系统均受影响,提出毒素说。
(一)枯萎病菌入侵棉株的途径
Bishop(1983)研究发现,在适宜条件下接种枯萎菌孢子24~96h后就能发生侵染,孢子萌发后,菌丝体在根表大量生长,主要从根尖分生区和伸长区侵入,很少侵入根冠、根毛。在茎尖的成熟区以上,由于组织发育成熟,病菌难以通过内皮层而进入维管束内。病菌主要是在成熟区之前或在寄主伤根时进入维管束。黄建成(1990)从植物解剖学研究了枯萎病菌入侵途径、病菌与各器官结构的关系之后提出,菌丝的侵入主要在苗期感染下胚轴及其根系,亦可见于子叶,在根、茎中菌丝主要定殖于最初形成的次生木质部。枯萎病菌在棉苗不同生育阶段的侵入途径是不同的。
1.棉苗期枯萎病菌的入侵
无菌培养的结果,病菌可在棉株任何部位侵入,但主要在下胚轴部位,在温室培养的棉株,常在土面以下5~10mm的下胚轴,此部位因受土壤温湿度、苗株与土壤摩擦等因素的影响,容易受损,使菌丝较易在此部位侵入。
下胚轴及根部的侵入。当菌丝接触下胚轴及其根系,常从破损的表皮细胞入侵滋生,并沿破损部位进入皮层,若菌丝比较多,也可从表皮细胞直接进入,表皮细胞无明显病变,菌丝进入皮层后,能在细胞内及细胞间生长,入侵的方法是径向的,菌丝常有粗、细的差异,粗的直径约为3.4μm,细的约为2.8μm,当菌丝进入中柱时,似乎不受内皮层的阻碍,若在下胚轴部位,菌丝沿着两个维管束之间的薄壁细胞到达髓部,然后向维管束的薄壁组织扩散,进入较大的孔纹及网纹导管,菌丝一旦进入维管组织的木质部,大体上就限制在较大的导管内,并向上扩展,以菌丝或分生孢子的形式,进入茎部。
子叶部位的侵入。从接种病菌后的幼苗,菌丝入侵子叶,当初菌丝先在表皮上滋生,细胞变褐,部分溃烂,并扩展到邻近细胞,其后细胞胶状破毁,菌丝沿破壁毁处,深入叶肉,菌丝可从细胞间及细胞内穿过。受侵染的叶片厚度变薄,与正常叶比较,约减少102μm。
2.现蕾前后的根、茎、叶解剖
根内的感染。从幼根的横切面看,菌丝可在表皮、皮层、维管组织中的薄壁组织中发现,并呈向心径向侵入,具有次生结构的根中,菌丝常限制在最初形成的次生木质部的导管中,它的纵向扩展则沿导管上升或下移,径向扩展是通过木质部射线细胞引伸。
从老根的横切面看,木质部射线感病尤为严重,其壁黑褐色,离心引伸,放射状排列,壁加厚,有时具有菌丝。与射线邻近的薄壁细胞及导管,常先感病。从老根纵切面看,菌丝多集中在导管内,呈分枝状,引伸方向大致与导管主轴平行,菌丝可由一个导管经过纹孔进入另一导管,有时可看到分生孢子的存在,个别导管可看见胶状物和侵填体,呈黄褐色,有时附在导管内壁上,侵填体呈囊状,在一个导管中往往数量较多,个别
茎内的感染。菌丝从根部侵入后,寄生在根部的导管及其邻近细胞内,随着导管向上输水,菌丝可向上、向下延伸,分生孢子也随水分上升而入侵茎、枝条、叶柄,并在这些部位的导管及其邻近细胞中萌发滋长。最初感染枯萎病的棉株,一般在茎秆外部形态上无特殊表现,稍后的生长,其节间比正常的缩短。从横断面可看到靠近髓部的木质部变黑褐色,若感染黄萎病,则可扩展到整个木质部,这是区别黄萎病及枯萎病的方法之一。
在现蕾期前后,进行病株茎的解剖,其菌丝的定殖部位,与根基本相同,即定殖于最初形成的次生木质部中,并有大量的孢子及菌丝存在,有些孢子在导管及其邻近的细胞内萌发,初期即可见菌丝从一个细胞伸入另一细胞。菌丝是不断向邻近细胞入侵,木质部的射线细胞往往感病严重,并存有孢子,可能与其横向感染有关。
叶的感染。棉花枯萎病在子叶期即表现病状,可分为黄色网纹型、紫红型、黄化型和急性青枯型,但以黄化型为基本性状,表现为叶子黄色兼有大块变色枯焦斑,最后叶子脱落。
苗期从一些叶的横切片中,可看到一些菌丝在栅栏组织及海绵组织中滋长,可在细胞内或细胞间穿行,具有菌丝的细胞,叶绿体相对减少,壁加厚,表皮细胞也受损。后来在该部位呈黄色网纹状及大块枯焦斑。
入秋后叶的横切面观察,叶柄的维管束可呈现黑色,细胞壁加厚,导管中可看见菌丝的存在,但不普遍,如具菌丝,多为扭曲或分枝状。
(二)导管堵塞
当枯萎病菌侵入棉株体内后,因菌丝及孢子的大量繁殖而堵塞了棉株的导管;或者刺激邻近的薄壁细胞产生凝胶体和树胶等胶状物质或侵填体(tylose)而填塞导管;也可能是病菌入侵后,产生果胶酶,使棉株细胞中的胶状物质和细胞壁中的果胶物质被水解,引起组织解体而堵塞导管。
导管被堵塞后,机械地阻碍了水分和养分在棉株体内的正常运输,加上地上部的蒸腾作用和呼吸作用旺盛,使水分失去平衡而导致棉株萎蔫。
袁红旭等(2002)观察到接种枯萎病菌后棉苗病株茎内大小导管内有数目不等的菌丝体。少的每个导管内1~2根菌丝体,多的可充满整个导管。病菌菌丝多沿导管纵向生长,也能看到菌丝穿过导管壁横向生长进入薄壁细胞或其他导管,导管周围的薄壁细胞中同样存在菌丝,但韧皮部中未发现菌丝体。被侵染的导管数目依发病情况变化而变化,发病严重的感病品种显著高于抗病品种。受侵导管及周围薄壁细胞常发生褐变,这种现象在导管有大量菌丝时很常见,但也能观察到变褐的维管束中并没有菌丝的现象。在病害发展的早期没有明显的褐变现象,只有在病害进一步发展、植株有明显的症状出现时,褐变才大量出现。引起褐变的物质不仅横向扩展而且可随导管汁液纵向扩展。发病棉苗茎部产生填充物充塞导管,这种现象所有品种都存在,未接种的对照中没有观察到填充物。填充物有两种类型:一种为球状物;另一种为胶状物,用酸性品红染色时胶状物为橘红色,球状物为鲜红色;用番红染色时胶状物为鲜红色,而球状物为淡红色;用苏丹Ⅲ染色胶状物为橘红色,球状物无色;用10%三氯化铁染色,两种侵填物均不显色。由此可见,两种侵填物成分不同,但都不是单宁类物质,胶状物可能是脂类。此外,病菌粗毒素诱导的棉苗维管束褐变和导管堵塞现象与发病植株维管束的变化情况基本相同。堵塞导管的填充物也有两种类型:球状和胶状。两种填充物对染料的颜色反应与发病植株中的两种填充物相同。毒素处理后,处理根苗叶柄也存在维管束褐变和导管堵塞反应,而且发生较茎部为早。但试验观测到病株内由菌丝及填充物堵塞的导管仅是一小部分,即使是发病严重的植株被堵塞的导管也只是总导管的1/10。黄建成(1990)的研究结果指出,在现蕾前期调查统计感枯萎病植株维管束的1879个导管中,含菌丝有104个,在此范围内,其他细胞具菌丝可达40个,但有些材料的其他细胞,不含菌丝,具有感病性状。感病的导管和其他细胞的壁均变黑、加厚,其厚度为5.67~8.5μm,而正常壁的厚度为3.4~5.6μm,仅增厚约在3.5μm;具侵填体的导管数占总导管数的0.68%,其比例较小,在未观察的材料略有增加,但增幅不大。这说明单纯的导管堵塞论并不能完全解释病株萎蔫过程。早在1954年,Waggoner的研究报告指出,木质部导管堵塞可以引起水分运输障碍,
(三)毒素致害
不少病原菌能产生可以扩散和转移的、对植物有毒的活性化合物,而导致对某些植物的病害。不少学者认为,棉花枯萎病菌入侵棉株后,引起导管的阻塞,只是导致棉株萎蔫的部分原因。更重要的是棉株体内产生某些有毒物质,使寄主细胞中毒死亡。
病原菌对寄主产生有毒物质,早在19世纪末便有人提出。其后,进行了广泛的研究,并取得了较大的进展。在棉花方面,Brand早在1919便指出,棉株枯萎性状的发生,除导管堵塞外,还可能由于植物组织中毒。据国内外报道,当枯萎病菌侵染棉株后,分泌出对寄主有毒的物质,主要有类萜、酚类、糖甙及其衍生物。有的病原菌能分泌出多种的多糖水解酶,不仅可使寄主细胞壁降解,细胞分离,菌丝得以乘机而入;而且使寄主根部受伤,吸水力下降。有的病原菌可分泌出两种水解酶——蛋白酶和酯酶,它们可分解寄主细胞的膜蛋白和膜脂类物质,使细胞膜透性增大,散失水分加快,而引起萎蔫。
棉花枯萎病菌的致病毒素是由枯萎病菌产生的镰孢菌酸(Fusaric acid,FA),又称萎蔫酸。它是一种非特异性的活体毒素(Vivotoxin),其学名为5-丁基吡啶-2羧酸(5-N-butylpiocolinic acid)(Keen,1972)。由于它的毒害作用,使寄主的保卫系统受到破坏,因而感病棉株的叶片出现网纹、皱缩或枯死;细胞原生质对水分的溶透性以及整个棉株的水分平衡受到破坏;多酚氧化物被抑制,从而产生明显的病理变化。Ganmann(1958)认为,萎蔫素一方面降低了细胞的保水能力,造成棉株失水;另一方面也破坏了原生质膜的渗透性,损坏了植株细胞膜的功能。
仇元等(1963)发现,用枯萎病菌的培养滤液处理棉苗可以导致萎蔫。张献龙等(1993)将鄂棉14品种的3~4叶期切根苗插入盛有不同培
品系镰刀菌酸培养滤液(%)处理后36h处理后72h处理后108h发病率(%)病指发病率(%)病指发病率(%)病指抗感抗感抗感抗感抗感抗感新陆早16号2531.641.77.914.610.525.02.610.410.533.36.618.85081.8100.031.845.891.7100.061.768.6100.0100.065.979.6100100.0100.039.670.8100.0100.072.975.0100.0100.095.8100.00(ck)0.00.00.00.00.00.00.00.00.00.00.00.0新海21号258.333.32.116.78.348.32.125.016.751.74.225.05081.890.022.737.5100.0100.047.760.0100.0100.047.779.6100.075.0100.027.152.1100.0100.070.875.0100.0100.091.7100.00(ck)0.00.00.00.00.00.00.00.00.00.00.00.0
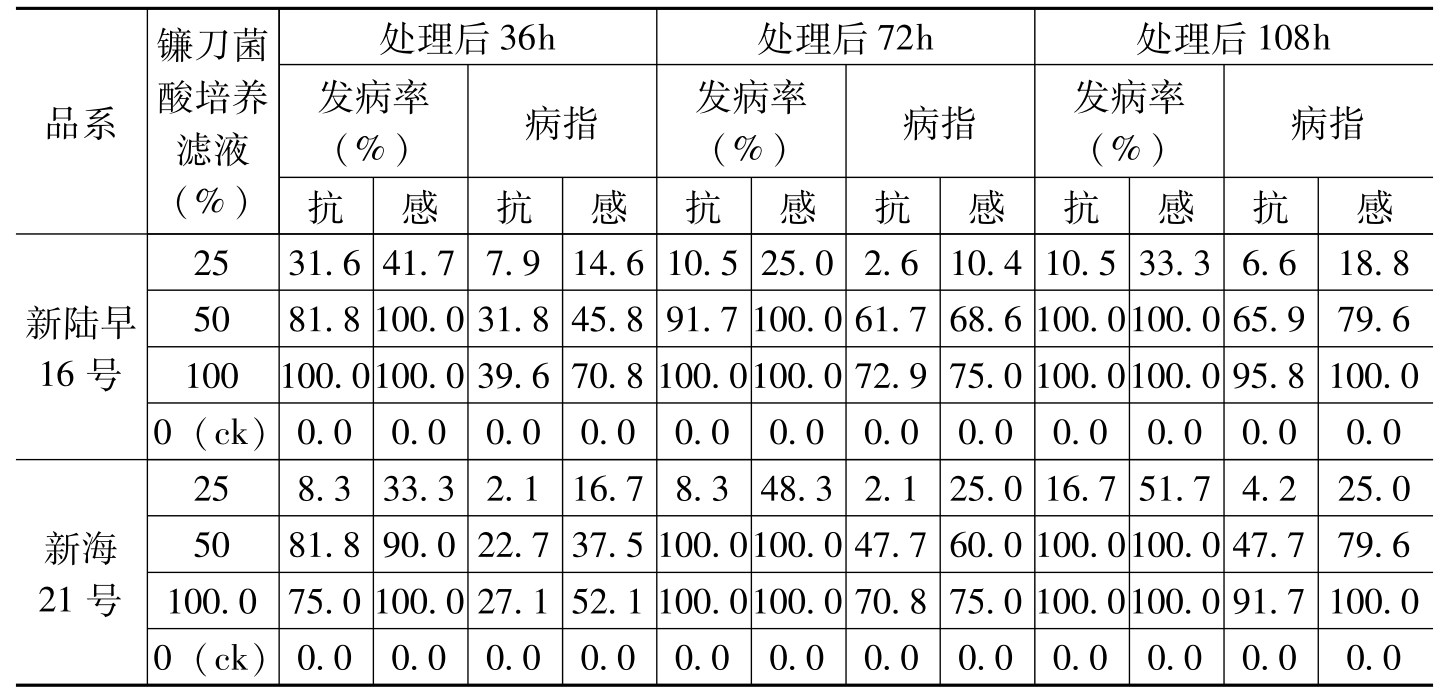
表3-1 病原菌培养滤液对新陆早16号和新海21号不同抗感品系的致萎作用
镰孢菌酸除可增加植物细胞电解质的渗漏、改变细胞壁的透性外,还能和铁、铜、锰等金属离子螯合,造成植物对可被利用元素的缺乏(Wood,1972;Wilson等,1978;Barna等,1983)。也有的研究表明,镰孢菌酸还能降低光合作用效率,抑制琥珀酸氧化酶及线粒体中细胞色素氧化酶,破坏植物体中的碳、氮代谢等。
所以,不少学者认为,枯萎病菌产生的镰孢菌酸的数量和致病力的强弱呈正相关。Chakrabarti(1979)用一个致病力强的菌株和一个致病力中等的菌株进行比较时发现,致病力强的菌株所产生的镰孢菌酸比致病力中等的菌株所产生的镰孢菌酸多2倍。当然,也有学者认为,病原菌致病力的强弱与各菌种产生的镰孢菌酸的数量没有直接关系。王贺祥(1984)用来自全国各地主产棉区的30个枯萎镰孢菌株分析指出,各菌株所产生的镰孢菌酸的数量,没有规律性;各生理型与镰孢菌酸的产量间也无相关性。
枯萎病菌分泌的酶类是否参与萎蔫病害的致病过程,多年来一直存在争论。Kumar等(1979)对棉花枯萎病菌在体外及接种后棉花植株中各种果胶酶的变化做了研究发现,病菌毒力与所产生的endo-PG(内切多聚半乳糖醛酸酶)量有相关性,无毒力菌株体外培养时和未接种的健康棉花植
为进一步肯定或否定endo-PG对棉花枯萎致病的作用,郜会荣(1990)利用紫外诱变方法获得了枯萎病的内切多聚半乳糖醛酸酶(Endo-PG)的缺失突变株(PGmⅠ为endo-PG完全缺陷菌株,而PGmⅡ菌株还有微量的endo-PG产生,但它们和母菌株相比差别是十分显著的),用突变株和野生菌株接种棉苗,发现突变株的致萎能力远远低于野生菌株。两周棉苗用母菌株接种时,快速萎蔫的棉苗可达26%~70%,用PGmⅠ接种的只有2%~4%,用母菌株接种,两天根表就满布菌丝,根表变褐色,根下部的维管束以外的组织发生软化烂掉,这种严重损伤的根可达40%~100%;PGmⅠ接种的棉苗,2~3天后根部也能布满菌丝,但根表变褐且颜色浅,3~4天后即有大量新的不定根长出,这些新发出的白色新根,发生软化和腐烂的根仅占6%~10%;PGmⅡ接种后,棉苗根部变化与PGmⅠ接种的相似,但发生软化或腐烂的比例较高,达10%~20%。在测定的病指方面,PGmⅠ接种后的病指高于PGmⅡ接种的,但这种差异在统计学分析上并不显著,而这两个突变菌株的病指都显著低于母菌株(表3-2)。可以看出,突变菌株PGmⅠ产生endo-PG能力完全丧失,致病力也大大降低;PGmⅡ菌株虽有微弱的endo-PG活力,而这种微弱的endo-PG活性可能在一定程度上对病菌的致病过程起一定作用,但这微弱的酶活性却不足以使菌株像母菌株接种那样达到高的病指。说明endo-PG参与了枯萎病菌的致病过程。Endo-PG主要通过降解植物细胞壁的果胶聚合物,使果胶胶体物质堵塞导管,影响棉株的水分运输,造成萎蔫。棉花枯萎菌的致病过程是酶和毒素协同作用的结果。
菌株病指重复A重复B平均病指差异显著性*母菌株53.656.054.8aPGmⅡ22.419.621.0bPGmⅠ8.816.012.4bc
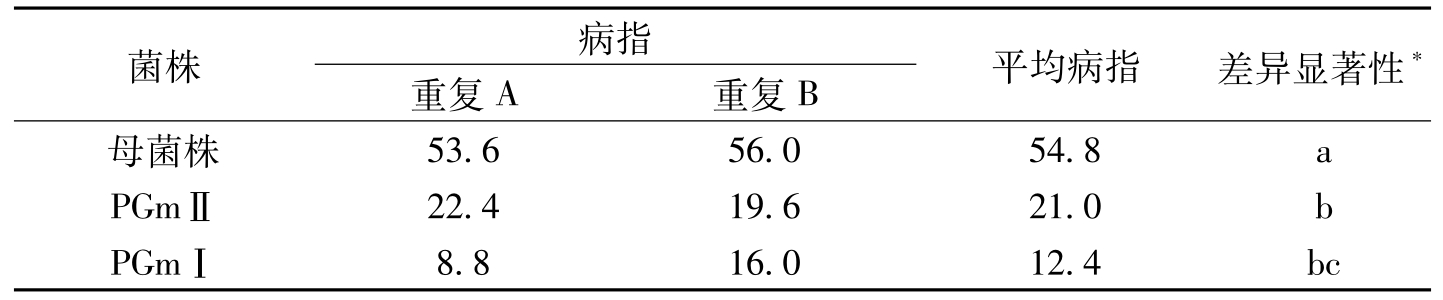
表3-2 各菌株接种棉苗一个月后病指的比较
由于寄主与病原菌之间的关系极为复杂,综合因素构成了棉花的凋萎。水分运输被阻塞或枯萎病原菌产生某种毒素可能只是导致棉株萎蔫的主要原因,有些因素的作用尚有待于进一步研究证实。
二、抗病机制
棉花对枯萎病菌的抗性是一个非常复杂的系统性问题。它涉及寄主(棉花)、病原菌、环境以及它们各自内部的生理代谢变化和相互作用。就寄主来讲,其抗病性可以分为组织结构抗性、生理生化抗性和生态抗性3个方面。但这3个方面的抗性又都可以归结到抗病基因上,即无论是组织结构抗性还是生理生化抗性和生态抗性都是由寄主的抗病基因所决定的。组织结构抗性是以物理的屏障阻止病菌的侵入和扩展。生化抗性是以植株体内所固有的或受到病菌侵染后新合成的抗生性化学物质对病菌的抑制作用。生态抗病性是指棉花根系所处土壤环境中,微生物和棉花根系分泌物对枯萎病菌侵染的影响。这3种类型的抗病性综合起来就决定了棉花是抗病的、耐病的、还是感病的。
(一)组织结构抗性
寄主植株的器官组织结构在抗病性中的作用占有重要的位置。若根、茎表皮细胞厚,根、茎木质部结构坚实,导管腔、木质纤维素腔直径较大,并且有多列髓和较厚细胞壁的棉花品种,均不利于枯萎病菌的侵入。所有品种的根、茎结构都是相同的,均具有初生保护组织的表皮,形状较大。排列较紧密,位于表皮与中柱之间的皮层,由维管束和髓、髓射线组
前苏联的一些学者研究认为,抗病品种基部具有坚实的木质部和含有大量淀粉贮藏物的多列髓射线,同时木质部的细胞间隙较小,细胞壁较厚。Bugbee(1970)的研究也认为,抗性品系的木质部导管比感病品系形成的多。对髓射线的研究各学者的意见较统一,即髓射线越多抗萎蔫病越强。这是因为在髓射线细胞中有一类称为类黄酮贮藏细胞(Flavarol-storing cell)的特殊细胞,这些细胞散布在髓线中,能阻止菌丝在各导管之间的扩展(Mace等,1981)。王正芬(1984)报道,棉花在6~7片真叶期,枯萎病的病指与茎导管细胞数之间有较强的正相关性。但对3~4片真叶期幼苗的研究却表明,病指与导管细胞数量的相关性很差。贺运春(1984)观察到棉株导管中的枯萎病菌菌丝壁由胞壁和胞膜两层组成,其厚度分别为0.14μm和0.10μm。枯萎病菌菌丝沿寄主导管壁生长时,菌丝细胞壁与导管壁紧密接触,在接触的菌丝细胞壁上产生顶端膨大、扁平、基部粗壮、不具有细胞壁的吸胞,并伸入寄主导管壁内。在菌丝的同一部位有时可以产生并排的两个相同的吸泡。在产生吸泡的菌丝部位,菌丝细胞加厚,厚度可达0.22μm。棉株导管中的枯萎菌丝无色,有分隔和分枝,并着生有小型分生孢子,但未见大型分生孢子及厚垣孢子。
Shi等(1992)利用组织化学方法,检测了继发性受阻导管和相邻不受阻导管的交联细胞的超微结构后发现,表现出细胞质成分及活性的增加,接触细胞能够产生脂类物质及其他化合物,所形成的分泌物通过纹孔进入导管。分泌物覆盖导管壁,并且以变态物形式沉积在导管腔,无定形泡状结构聚合在一起,形状、大小不一,导管逐渐被分泌物所包埋覆盖,累积的分泌物完全堵塞了导管腔。在抗病品种中,随着导管腔的分泌物积累以及管壁的增厚,被膜更集中,抗病品种中接触细胞中这种更集中的分泌物活性导致了屏障的形成,从而阻止了导管内病菌的扩展。雷江荣等(2010)利用农杆菌介导法将克隆自拟南芥的抗病基因
图3-1 棉株根茎木质部石蜡横向切片(胼胝质)
图3-2 棉株根茎木质部导管石蜡切片(×255)(侵填体)
维管束组织以外的细胞与抗枯萎病性也有一定的关系。史金瑞(1986)对枯萎病抗、感棉花品种根部早期侵染的组织和细胞进行观察发现,侵染菌丝在高抗的亚洲棉品种(石系亚)上局限在表皮细胞及外围1~2层皮层细胞中,而陆地棉中植86-1(抗)、徐州142(感)和高感的海岛棉品种中,菌丝能在皮层中扩展,并能进入中柱。研究中还发现,亚洲棉石系亚的根部表皮层外有一层物质,在菌丝与表皮细胞接触处这层物质被消解之后才能进行侵染,而在其他几个供试的陆地棉、海岛棉中则无此现象。石系亚对棉枯萎病表现较强的抗病性,可能与其表皮层外这层物质有关。
(二)生理生化抗性
棉花抗枯萎病的生理生化机制是个比较复杂的问题,枯萎病菌侵染棉花后,棉株体内的生化物质和生理代谢均会发生变化,反映了棉株抗、感病性的内在原因。
1.酶类
抗病性是植物的遗传潜能,其表现受寄主与病原的相互作用和环境条件的共同影响。根据“基因对基因”假说,植物抗病性是植物本身所具有的抗性基因(Resistance Genes)和与之相对应的侵染病原物所具有的无毒基因(Avirulencegenes)结合时所表现出来的(Hammond等,1997)。不同的植株对同一病原物,同一植株对不同种的病原物可具有不同的抗病性(鲁明波等,1998;Heath,2000)。因此,植物不同的抗病性反映在植物生理上就表现出一系列复杂的生理生化变化,包括植物细胞内活性氧的积累与清除、抗病信号的产生与转导、防卫反应的表达与调控等。在这一复杂过程中,一些相关酶类起着很重要的调控作用,如超氧化物歧化酶(Superoxide dismutase,SOD),过氧化氢酶(Catalase,CAT)和过氧化物酶(Peroxidase,POD)等。许多研究都表明,植物病害的发生与这些酶活性变化有着密切关系,并且非亲和性互作(抗病反应)和亲和性互作(感病反应)两者在SOD等酶活性变化方面有着显著不同。近年来研究发现,植物受到病原物侵染后,与抗病性有关的一些主动防卫反应,包括细胞过敏
(1)活性氧清除酶类
近十几年来,活性氧(Reactive Oxygen Species,ROS)因在生物体内广泛存在并具有多种生理功能而引起人们的极大关注。通常所指的ROS仅指氧自由基,如超氧阴离子(O
ROS通常被认为是植物正常代谢过程中的有毒副产物,随着对其研究的深入,发现其在植物与病原物互作的防卫反应中具有重要作用。首先,ROS具有直接的抗微生物功能,其存在本身就可对病原菌造成伤害;其次,ROS还可以触发植物受侵染点的细胞死亡导致过敏性坏死反应(HR);再次,ROS还参与细胞壁木质化及富含羟脯酸的糖蛋白的交联,这有利于抵御病原菌侵染;另外,ROS很可能作为第二信使调控抗病相关基因的表达,并启动植物抗毒素合成基因的转录。但是,当植物体内ROS积累过多,就是ROS与其清除酶类之间的动态平衡被打破时,植物就会受到伤害(膜脂过氧化和膜差别透性丧失)。因此,植物在长期的进化过程中,在利用氧的同时也形成了一系列清除活性氧为害的机制,这些机制中有些是通过酶促反应实现的,有些则是由非酶物质如抗坏血酸、维生素E、β-胡萝卜素等来实现的。为了早日弄清植物的抗病机制,许多病理学家就活性氧清除酶活性的变化与植物抗病性之间的关系进行了大量的研究,但目前仍未得到一致结论。不过,研究不同抗病品种感病后棉花体内的防御酶活性变化,有利于探明棉花抗病性机制。
①POD 过氧化物酶(POD)及其同工酶在植物机体防御体系中起重要作用。POD不仅参与了木质素的聚合过程,也是细胞内重要的内源活性氧清除剂,因此,POD活性与植物抗病性有着密切的关系。目前,植物抗感病品种POD活性变化与抗病性间的关系观点不一。一种观点认为,植物感病后,抗感品种POD活性均升高,并且抗病品种POD活性比感病品种
在棉花上沈其益等(1978)报道,POD与棉花对枯萎病的抗性有关。田秀明等(1991)的研究结果表明,在人工接种枯萎菌后,抗病品种和感病品种POD的活性都显著加强,但感病品种POD同工酶的反应比抗病品种强烈得多,而且感病品种比抗病品种多1~2条酶带,且颜色较深。无论是抗病品种还是感病品种,单株病指越高,POD同工酶带数越多,颜色也越深。从海岛棉、亚洲棉和陆地棉三大棉种接菌后POD同工酶谱来看,也同样有以上趋势。
胡小月等(1993)指出,苗、蕾期,在未感病的情况下,抗、感病品种间POD酶带无显著差异;在感病情况下,蕾期不同抗性品种POD酶带数均增加,而且酶带颜色加深,说明病害发生后,抗、感品种酶活性均加强,植株体内代谢旺盛,但不同品种间POD酶带存在差异。在健株中,供试品种均具有P
不同抗枯萎病类型品种和同一品种内感病株和健株POD活性变化不尽一致(图3-4)。由图3-4可以看出,同一品种(系)内感病株和健株比较,健株叶片内POD的变化幅度很小(差值6个单位),曲线变化比较平稳,而感病株POD变化幅度较大(差值69个单位),曲线表现陡直。对于不同抗病类型的品种(系)来看,感病品种叶片内POD活性的变化幅度最大(差值64个单位),耐病品种次之(差值45个单位),抗病品种变幅最小(差值5个单位)。由此说明,品种的抗病性能越差,过氧化物酶活性的变化幅度也越大。
抗病基因

图3-3 不同抗性品种间健株与病株蕾期叶片POD酶带
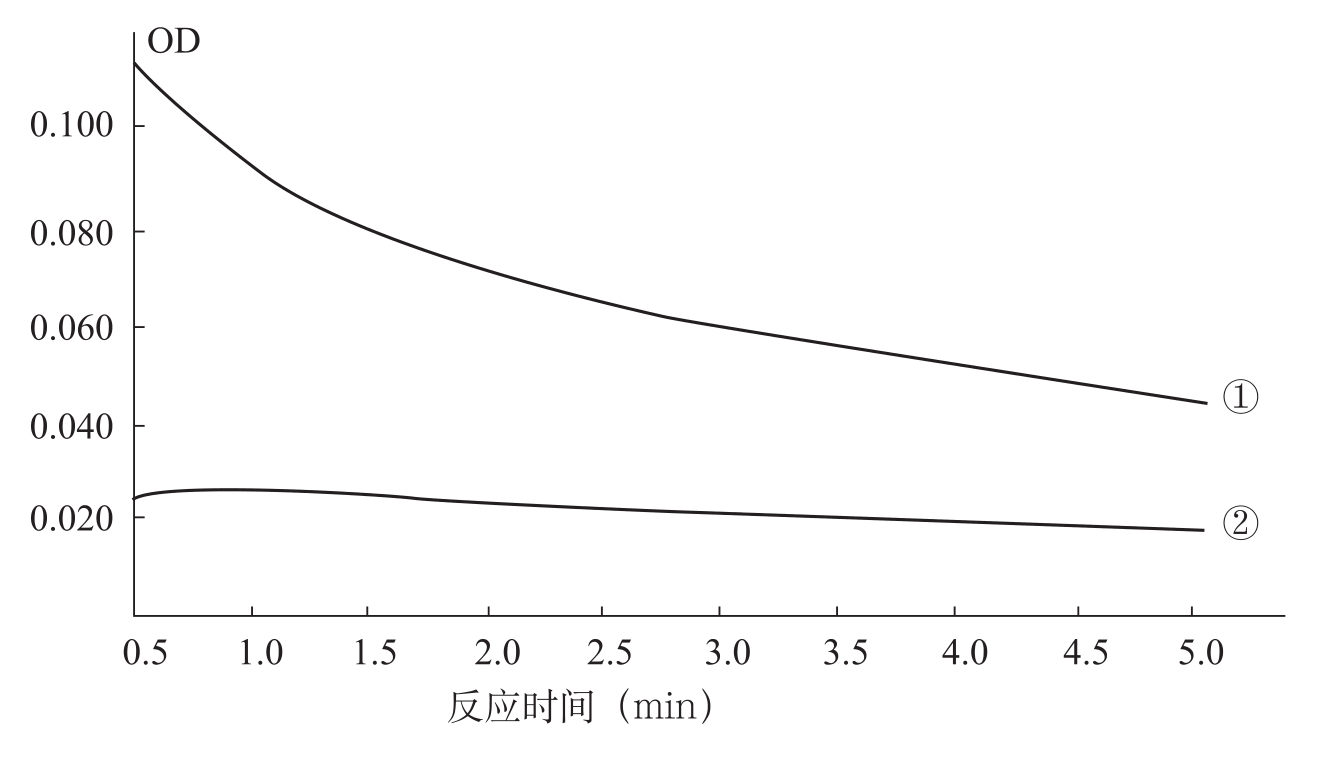
图3-4 同一品种感病株和健株POD活性变化(李妙等,1995)
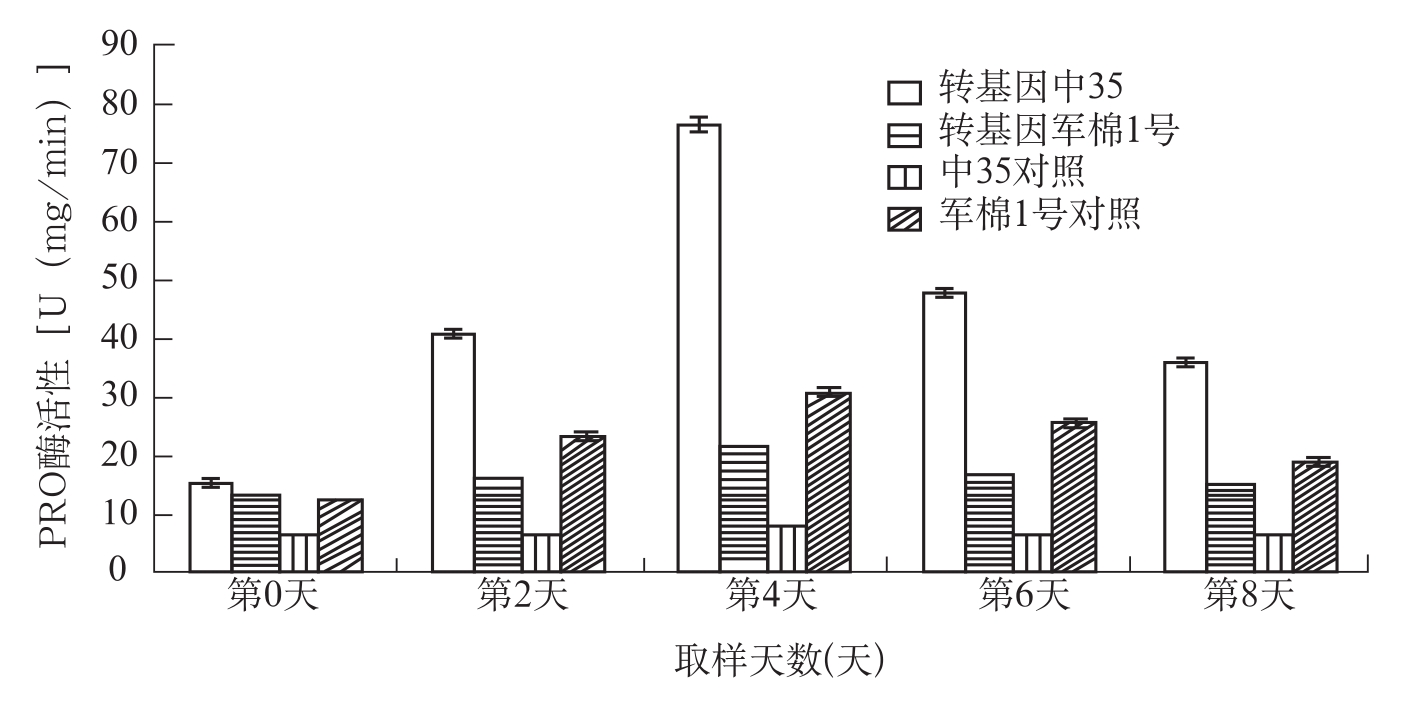
图3-5 棉株叶片接菌后过氧化物酶活性测定分析
POD广泛存在于植物体内的不同细胞定位,这些不同细胞定位的POD在植物抗病性中可能有不同的作用。有研究表明,受病原菌侵染或诱发物处理后,可导致植物叶片细胞间隙POD活性的增加,且与植物抗病性抗性有关。细胞壁中存在着丰富的POD,有人观察到在经诱发处理并产生有系统诱导抗性的植物叶片中结合于细胞壁上的POD活性明显上升,这些POD参与木质素在细胞壁上的沉积,从而与系统诱导抗性的产生相关。(Hammerschmidt等,1982;Ye等,1990)。宋凤鸣等(1997)观察到接种枯萎病菌后第8天的棉苗叶片和根茎部组织中的总可溶性POD、胞内POD和细胞间隙POD活性(表3-3)。由表3-3可知,未接种病菌的棉苗组织中3类可溶性POD活性抗病品种均高于感病品种,但根茎部组织中的总可溶性POD和胞内POD活性差异不明显;枯萎病菌侵染后,3类可溶性POD活性均有明显的提高,其中,胞间POD活性增加最大。这说明POD在棉花
品种处理过氧化物酶活性[△OD179/(mgpro·min)]叶片组织根茎组织总POD胞内POD胞间POD总POD胞内POD胞间POD中棉所12(抗病)不接种30.8121.3125.008.697.4818.47接种70.1332.14145.0038.0321.6439.806037(感病)不接种25.2716.3519.707.907.5010.89接种47.4434.6735.0028.2421.2624.26
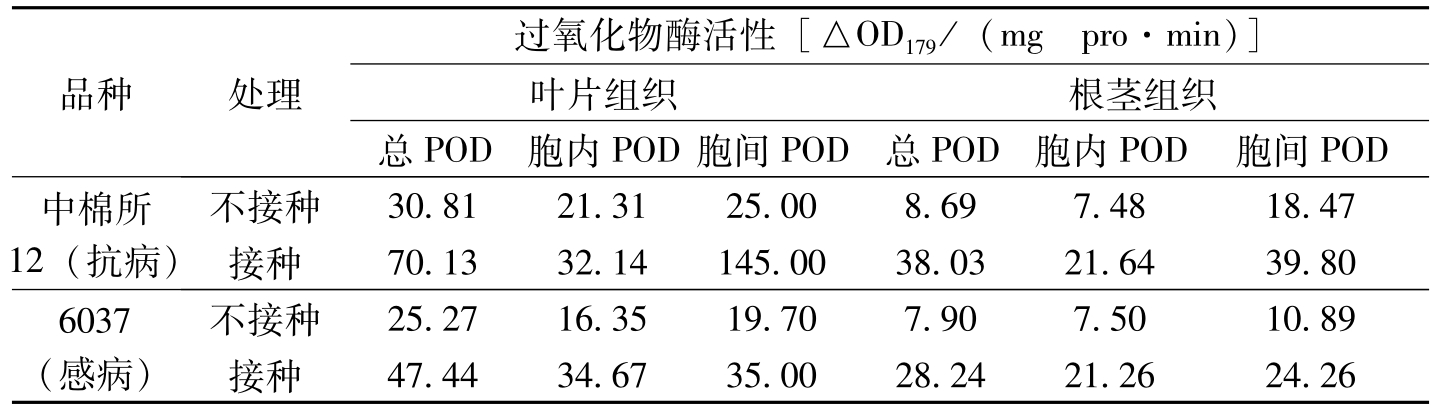
表3-3 枯萎病侵染后棉苗叶和根茎组织中总可溶性POD、胞内POD和细胞间隙POD活性
②SOD 超氧化物歧化酶(SOD)是植物细胞内防御酶系统的重要成员之一,它的生理作用是歧化O
在棉花上,吴小月等(1993)报道,超氧化物歧化酶在健株中均仅有3条酶带:P
在棉花发病高峰期,不同抗病类型品种中,感病类型SOD活性最高,耐病品种次之,抗病类型表现最低。这说明SOD活性与品种在田间感病的程度存在着内在相关关系(表3-4)。对SOD活性与田间病指进行相关分析得出,相关系数
图3-6 不同抗病品种间病株和健株蕾期叶片SOD酶带
抗病类型品种名称SOD活性[mg/(pro·min)]抗病中棉所1226.65石抗15524.17石30856.99石310423.79邯32212.77平均值18.87耐病冀8913535.13美861531.47美861639.16冀92.6134.66平均值35.11
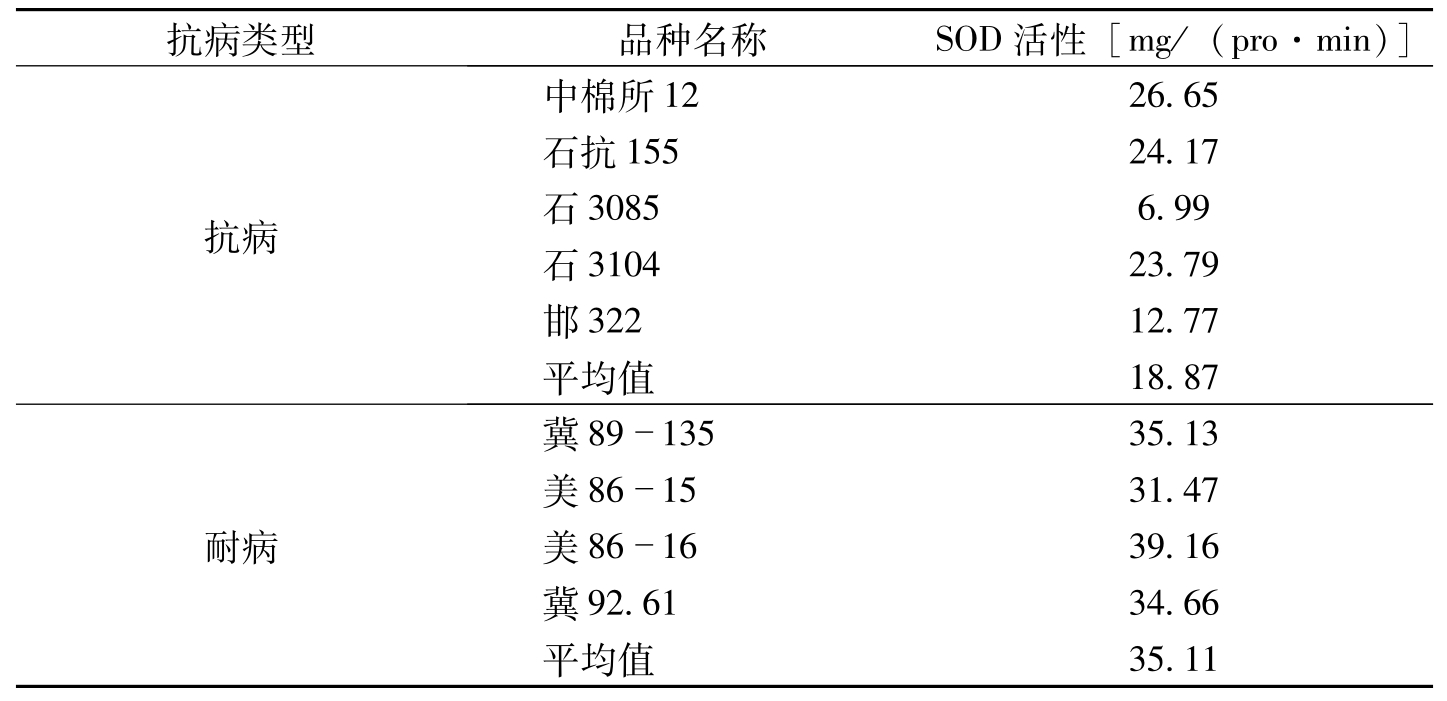
表3-4 不同抗病类型棉花品种(系)的SOD指标比较(李妙等,1995)
抗病类型品种名称SOD活性[mg/(pro·min)]感病邯89s9347.43冀92.10363.22选148142.83选155148.35平均值50.46
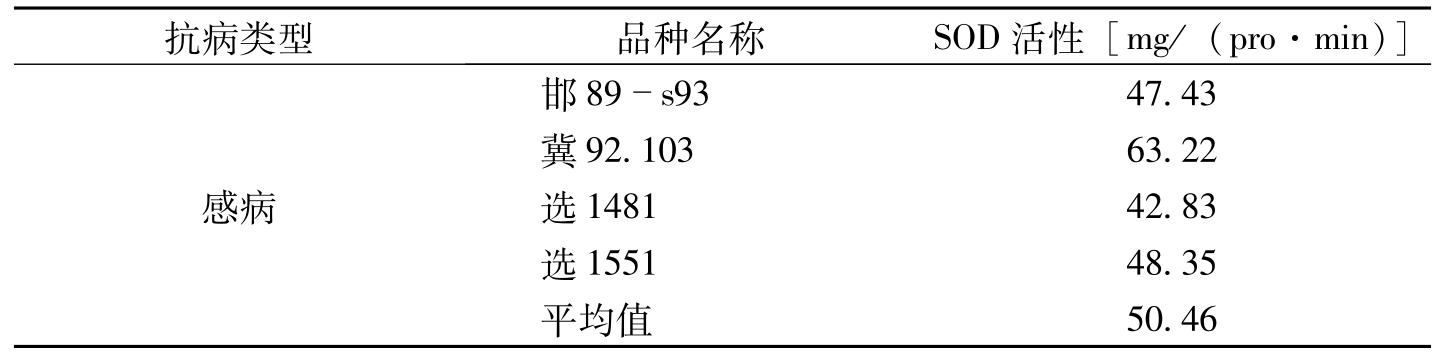
表3-4 不同抗病类型棉花品种(系)的SOD指标比较(李妙等,1995)(续)-1
受病原物侵染后植物体内SOD等保护酶活性的变化在亲和性互作引起的感病反应中SOD酶活性升高,且与病害症状的表现有关,但在非亲和性互作引起的抗病反应中,SOD酶活性无明显变化甚至下降。宋凤鸣等(1999)研究结果指出,健康棉苗中,抗病品种和感病品种间SOD酶活性无明显差异,试验期间酶活性无显著变化;枯萎病菌接种后棉苗组织中SOD酶活性明显升高。感病品种6037在接种病菌后3天时SOD酶活性就显著高于对照,此后呈直线上升,而抗病中棉所12的SOD酶活性在接种病菌后7天才开始上升。雷江荣等(2010)报道,转
植物受病菌侵染后可选择性地刺激SOD同工酶活性的变化,如亲和性锈菌侵染后菜豆组织中Mn-SOD酶活性大幅度增加,而非亲和性锈菌侵染后Cu、Zn-SOD酶活性明显提高(Montalbin等,1986)。受TMV侵染的烟草叶片中SOD酶活性的增加主要来自于Cu,Zn-SOD(Buonario等,1987)。宋凤鸣等(1999)研究的结果证明,枯萎病菌侵染后棉苗组织中POD酶活性的升高主要是由Cu、Zn-SOD活性的增加引起的(表3-5,表3-6)。
叶片组织根茎部组织SOD活性[U/(mg·min)]抑制率(%)SOD活性[U/(mg·min)]抑制率(%)酶粗提液75.4080.63KCN0.01mmol/L74.601.0079.431.490.10mmol/L60.6819.5271.9019.950.20mmol/L40.4840.3162.5022.490.50mmol/L15.3779.0242.1747.701.00mmol/L6.2891.0719.4675.87氯仿乙醇(1∶2)70.935.9067.3416.48
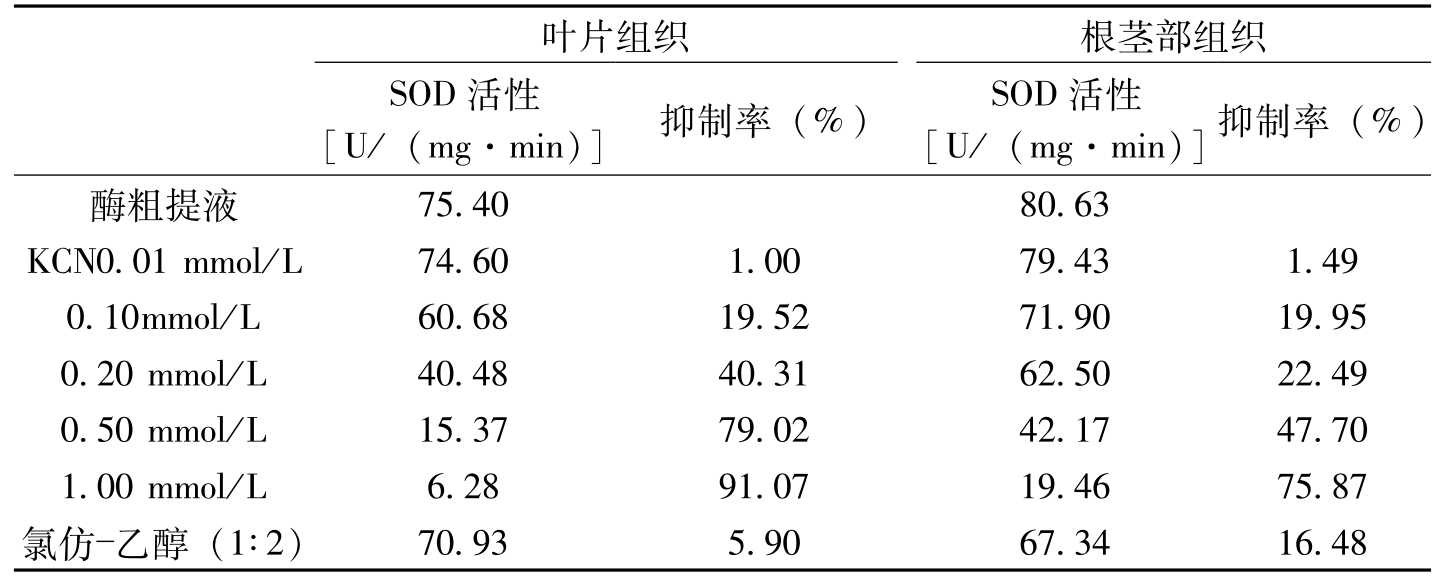
表3-5 KCN和氯仿对棉花SOD活性的抑制
[U/(mg·min)]品种处理叶片组织根茎部组织总SODCu,ZnSODMnSOD总SODCu,ZnSODMnSOD中棉所12(抗病)不接种35.1833.281.9020.0017.202.80接种39.7037.302.992.8322.835.006037(感病)不接种30.8834.272.3124.2220.903.45接种60.8956.674.2850.2743.656.95
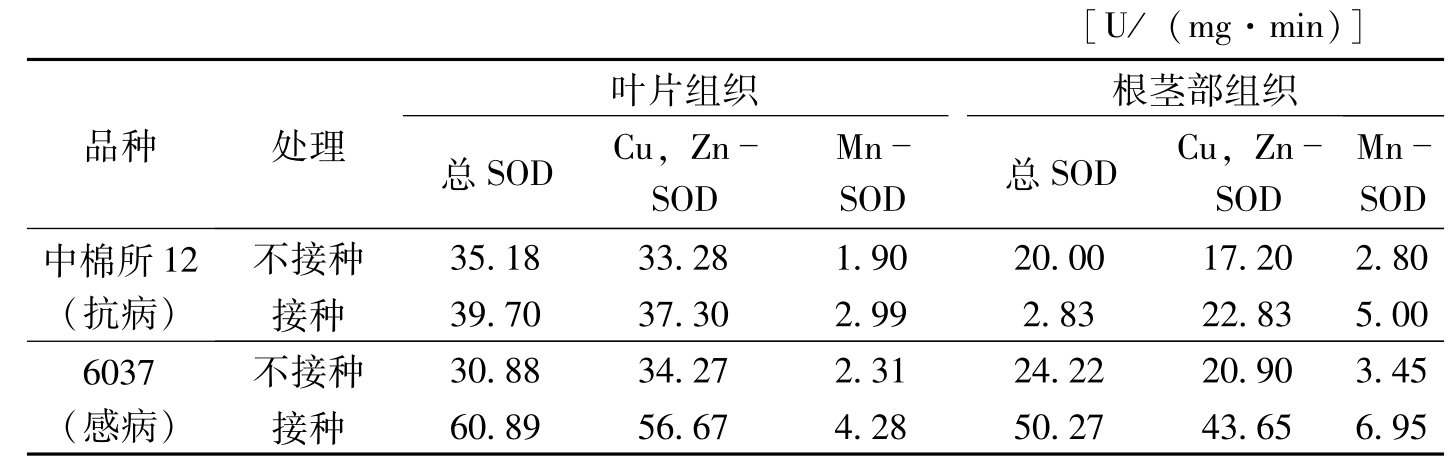
表3-6 枯萎病菌接种后7天棉苗组织中总SOD,Cu,Zn-SOD和Mn-SON活性
③CAT 在多种植物—病原物互作系统中都有H
在棉花上,宋凤鸣等(1999)报道,健康棉苗中,两个供试品种组织中CAT酶活性无明显差异;接种枯萎病菌后,棉苗组织中CAT酶活性显著增加,与抗病品种中棉所12相比,感病品种6037的CAT酶活性上升早而显著。
雷江荣等(2010)的研究结果指出,采用农杆菌介导法将来自拟南芥的抗病基因
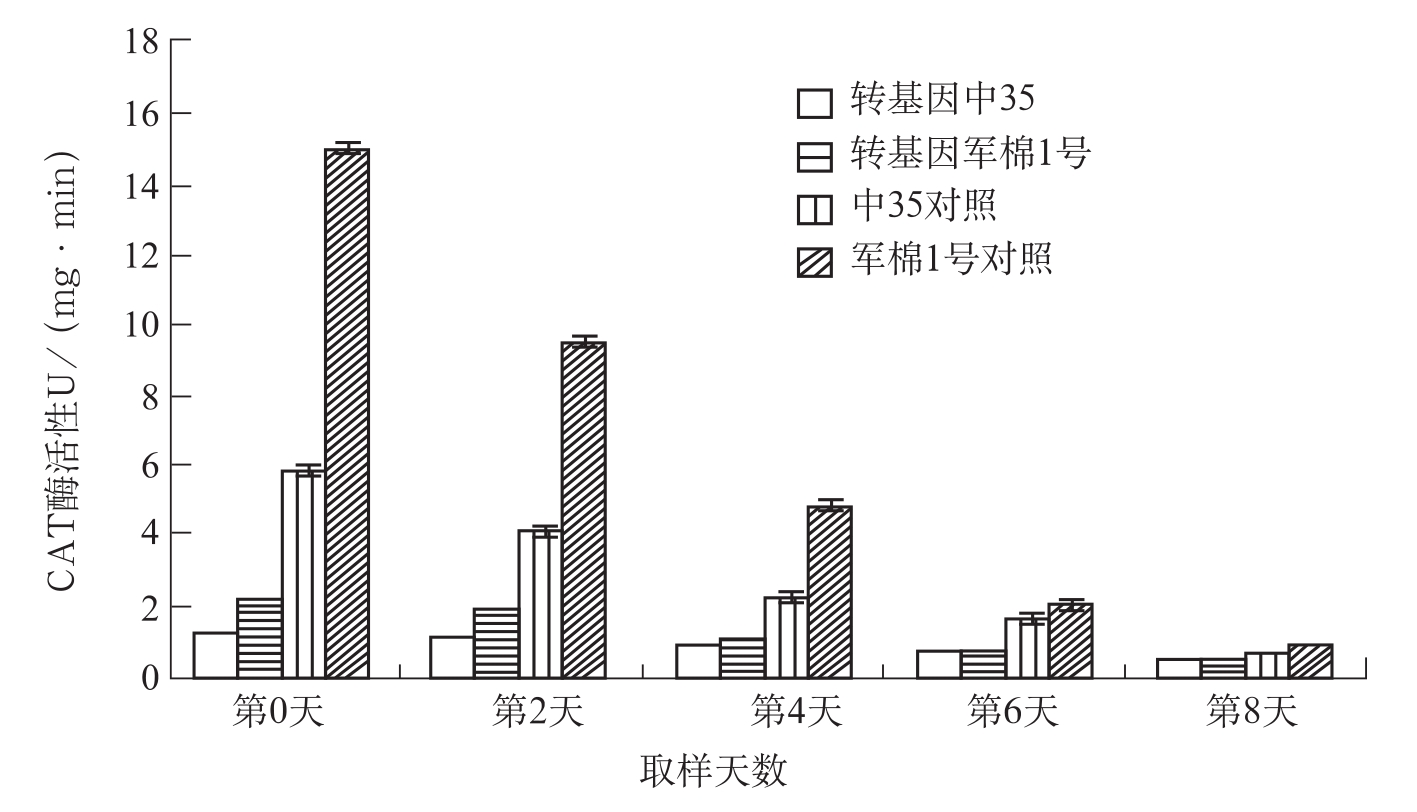
图3-7 棉株叶片接菌后过氧化氢酶活性测定分析
第0天第2天第4天第6天第8天转基因中351.167±0.0291.012±0.0250.868±0.0210.663±0.0160.415±0.010转基因军棉1号2.1217±0.0531.835±0.0451.061±0.0260.645±0.0160.407±0.010
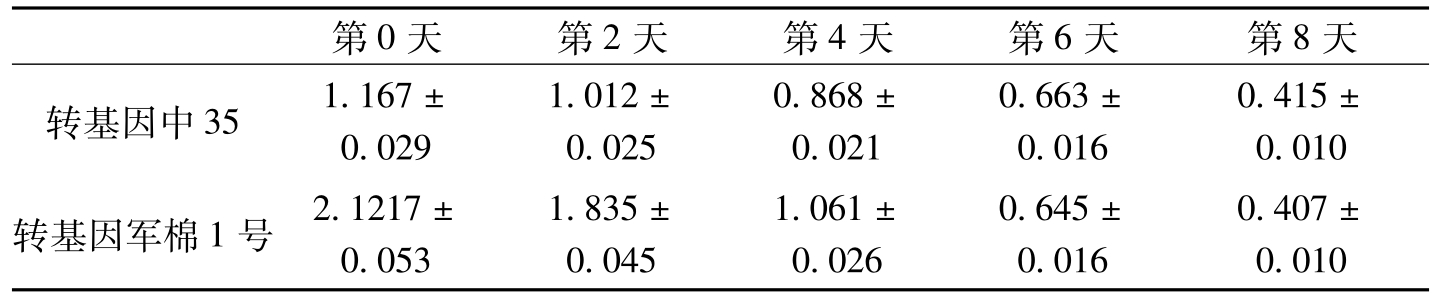
表3-7 棉株叶片接菌后过氧化氢酶活性测定分析
第0天第2天第4天第6天第8天中35对照5.769±0.1444.086±0.1022.188±0.0541.597±0.0390.611±0.015军棉1号对照14.989±0.3749.504±0.2374.820±0.1201.946±0.0480.866±0.021
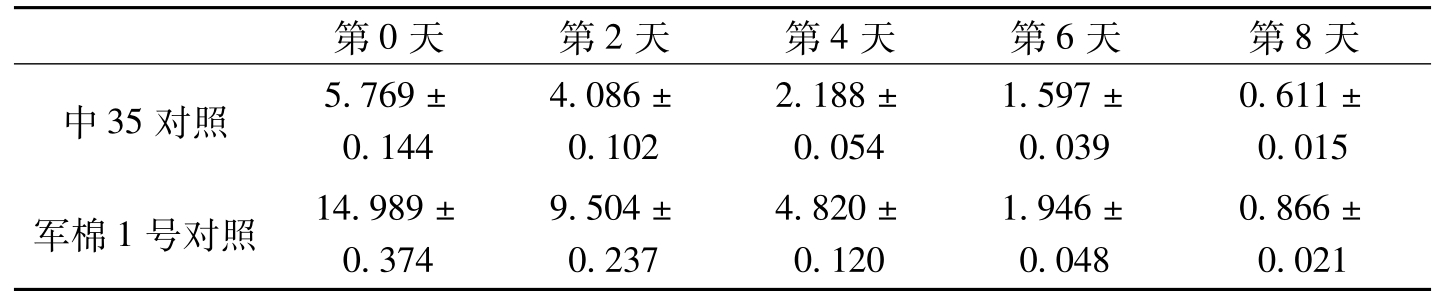
表3-7 棉株叶片接菌后过氧化氢酶活性测定分析(续)-1
(2)抗病反应次生代谢酶类
植物次生代谢产物(Secondary Metabolites)是指植物体中一大类并非生长发育所必需的小分子有机化合物,其产生和分布通常具有种属、器官、组织和生长发育的特异性;抗生作用是植物次生代谢的一个重要生理功能,也可视为自然选择的结果,参与植物防御的次生物质很多,包括酚类、植保素、木质素和其他一些次生代谢物。与这些次生代谢有关的主要酶类是多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)。
①PAL PAL是植物苯丙烷类次生代谢途径总路第一步关键酶,是苯丙烷类代谢途径的关键酶和限速酶,它催化苯丙氨酸脱氨基后产生肉桂酸并最终转化为木质素,因此,它是与细胞内木质素生成和沉积有关的防御酶。当病菌入侵时,细胞受到刺激后启动PAL系统产生木质素并沉积在细胞壁周围,将病原物限制在一定的细胞范围内阻止其进一步扩散为害。自从1964年Minamiawkd和Uritain首次发现植物感病后PAL活性增强以来,陆续有许多研究证明植物受到不同病原体感染后PAL活性均有升高的现象。因为PAL活性的升高往往与植保素、木质素等抗性物质的产生和积累呈正相关。王敬之等(1982)提出,把PAL活性作为植物抗病的生理指标加以研究。
在棉花上,冯洁等(1990)研究了棉花抗、感品种感染枯萎病后体内PAL的动态变化。结果表明,在根内,无论抗、感品种接菌后的PAL活性都始终高于未接菌的对照。抗病品种86-1、陕1155在接菌后24h,经病原菌的诱导出现一个酶活性峰,而感病品种在接菌后36h才出现PAL活性峰,比抗病品种晚12h。抗病品种的相对酶活值也明显高于感病品种,其幅度是感病品种的1.38~2倍(图3-8)。在叶片内,抗病品种在接种枯萎菌后分别于12h和24h出现了PAL活性峰(86-1在接种后36h又出现
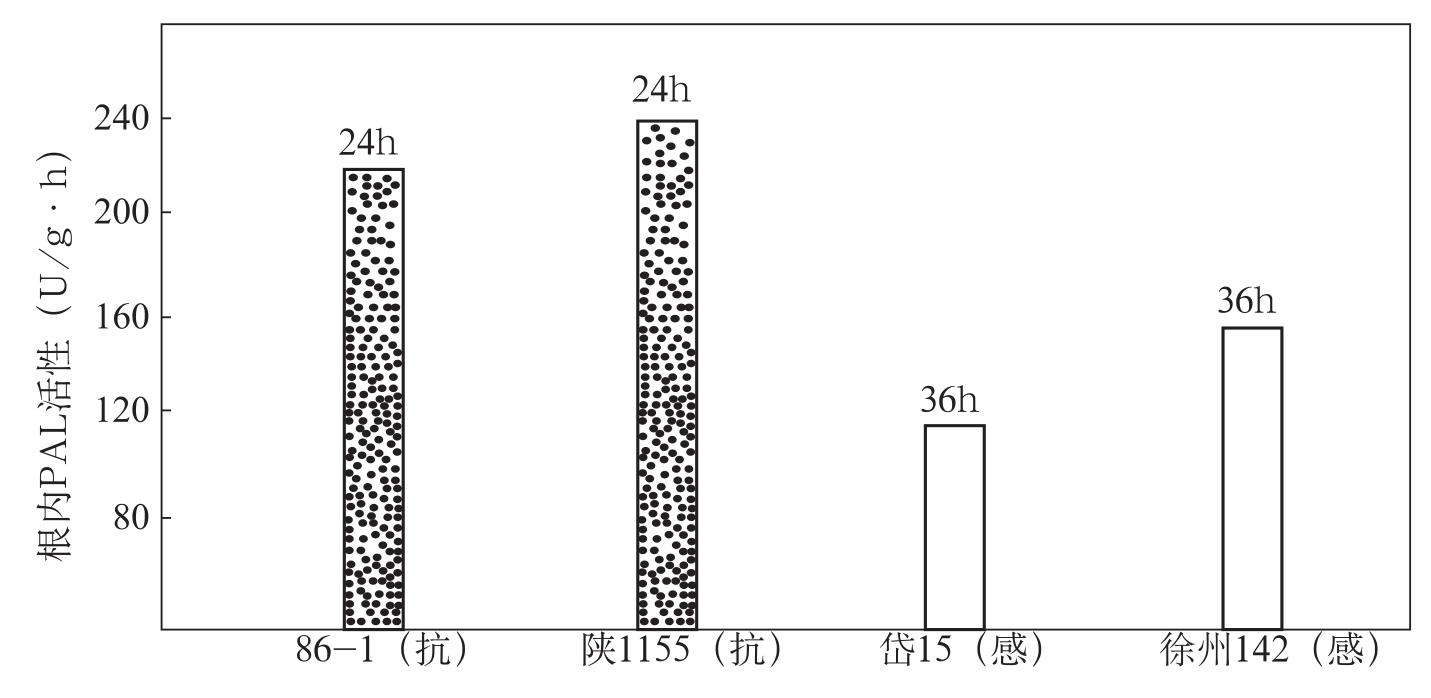
图3-8 抗、感品种接菌后根内PAL活性峰的相对酶活值
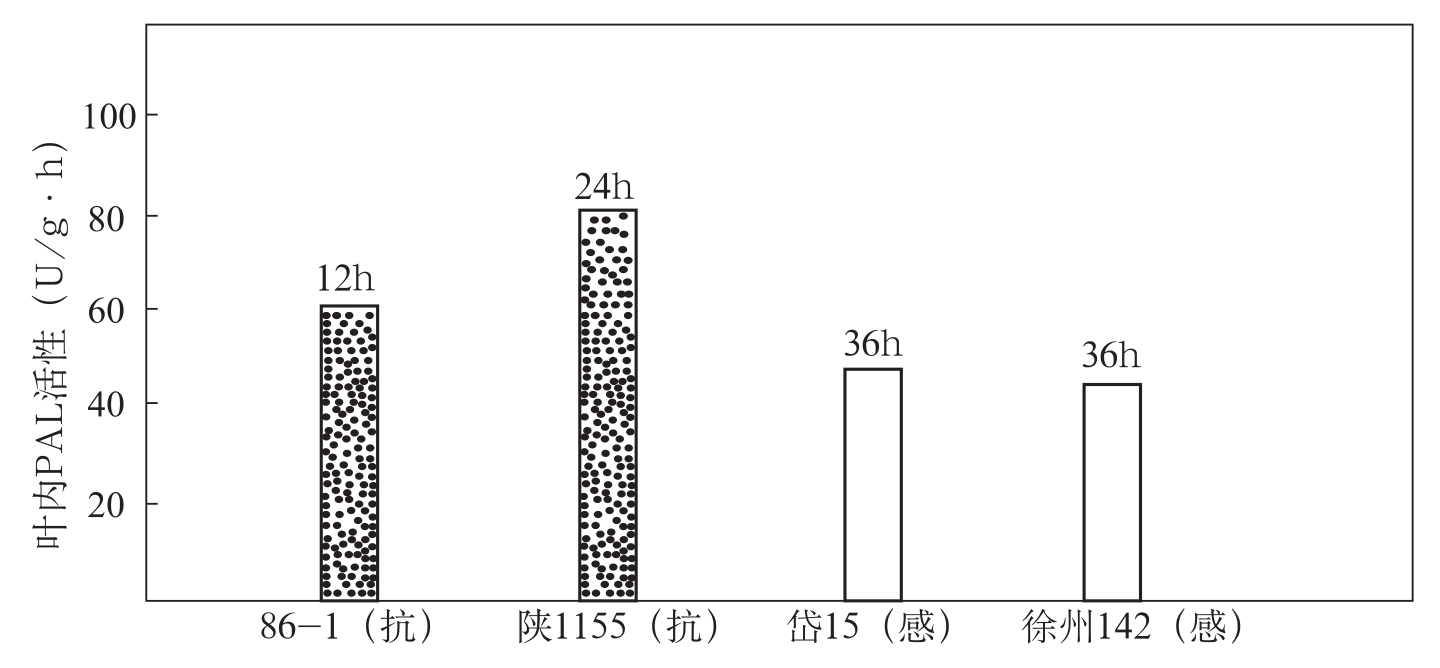
图3-9 抗、感品种接菌后叶内PAL活动峰的相对酶活值
袁章虎等(1995)报道,接种枯萎病菌后抗病品种的PAL活性迅速提高,并很快在24h到最高峰。而感病品种的PAL活性则相对上升较慢,在36h才达到最高峰。其峰值也比抗病品种低得多,前者最高峰值为5.68U/(g·h),而后者仅4.12 U/(g·h)。这说明品种的抗病性不仅
转抗病基因
总之,棉株在受到枯萎菌侵害后能够在很短的时间内迅速提高PAL活性,从而合成较多的抗生性物质,抑制病菌在棉株体内的萌发和扩散,使棉株表现抗病性。反之,如果病菌顺利地在寄主体内萌发、定殖和扩散,就使棉株表现感病。
[U/(g·h)]品种第0天第2天第2天第2天第2天转基因中353.485±0.0874.685±0.1176.007±0.1515.837±0.1454.835±0.120转基因军棉1号2.072±0.0513.887±0.0974.723±0.1184.091±0.1023.905±0.097中35(对照)2.838±0.0703.255±0.0813.885±0.0974.011±0.1003.778±0.094军棉1号(对照)1.928±0.0482.005±0.0502.394±0.0592.622±0.0642.210±0.055
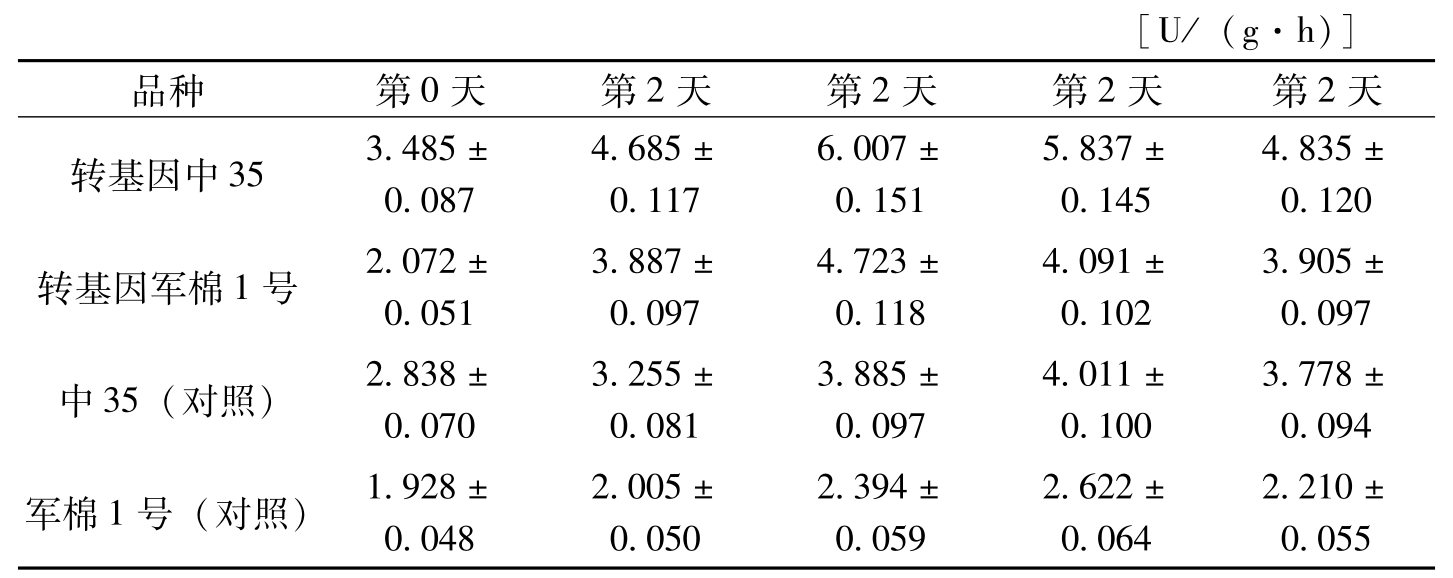
表3-8 棉株叶片接菌后苯丙氨酸解氨酶活性测定分析
大量研究表明,多酚氧化酶(PPO)主要参与酚类氧化为醌以及木质素前体的聚合作用,与植物抗病密切相关。病原菌侵染能诱导植物体内PPO活性升高,促进酚类化合物在受侵染部位的合成和积累,大量的酚可由多酚氧化酶氧化成醌,醌类化合物能钝化病原物的呼吸酶,阻碍病原物的生长,醌的次生反应所产生的黑色素的痂可阻止感染的扩散;酚类化合物是细胞形成木质素的前体,可形成木质素,促进细胞壁和组织的木质化,以抵抗病原的侵染。
在人工接种枯萎病菌后,感病品种PPO活性的初期受到一定的抑制。直到48~72h才超过对照,而抗病品种PPO活性从24h就明显增强并超过
在研究转抗病基因
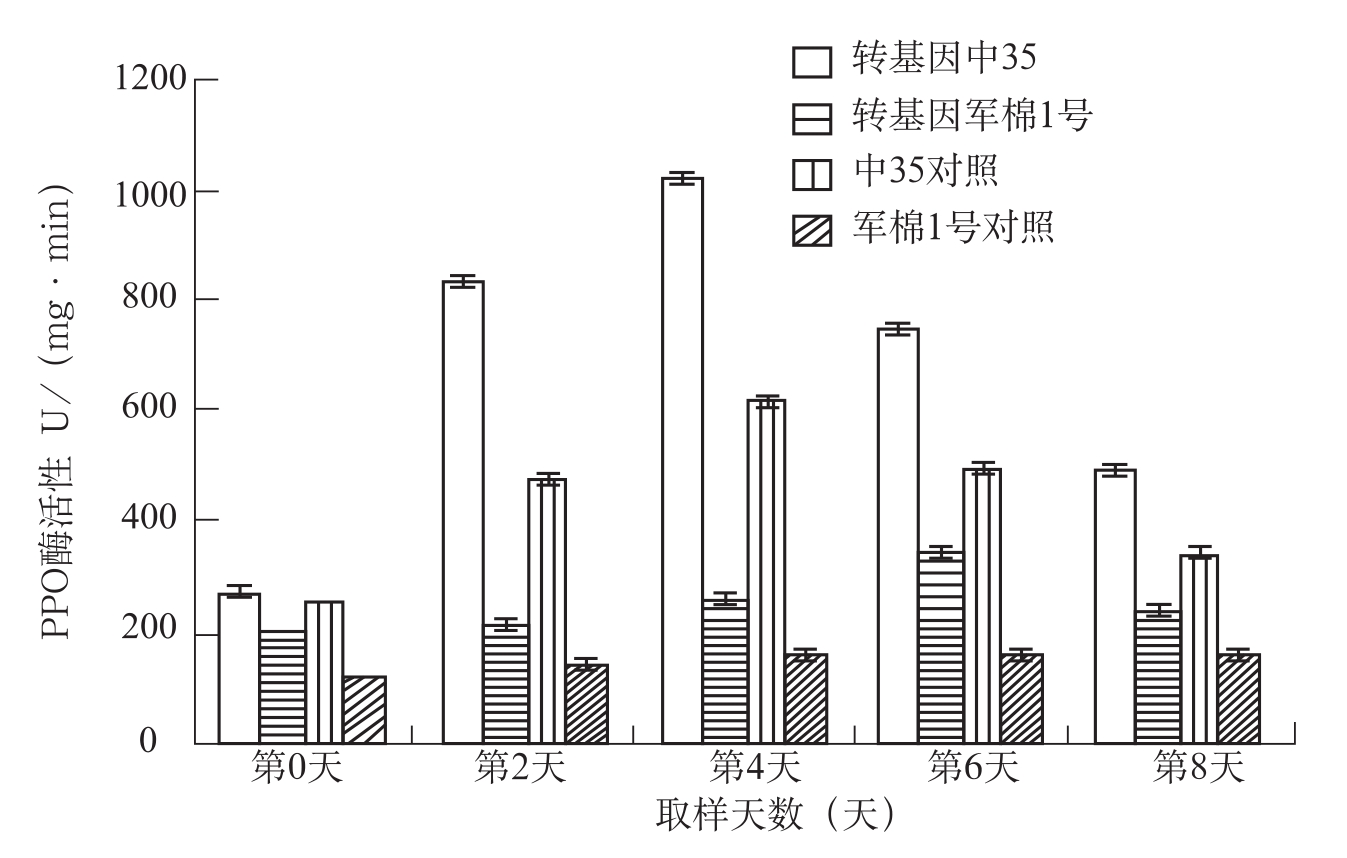
图3-10 棉株叶片接枯萎病菌后多酚氧化物酶活性测定分析
从总体上看,抗病品种和感病品种在接种枯萎病菌后都相继出现不同程度的PPO活性高峰。其区别是抗病品种出现的早,峰值高,而感病品种出现的晚,峰值低,抗病品种的PPO活性不受病菌的抑制,而感病品种则在受侵染的初期受到一定的抑制。
研究棉花感病后防御酶活性变化,有助于深入研究棉花抗病机理,为
2.糖类
在染病的植物中,糖不仅是植物各类代谢的基础,而且也可以作为病菌的营养。糖代谢能为蛋白质、脂肪、核酸及次生代谢提供碳骨架和能量来源。陈其煐等(1990)以采自河南、湖北、辽宁、江苏、新疆等棉区棉花枯萎菌对具抗枯萎病性的陆地棉品种86-1、陕1155,感病品种岱15、徐州142和鲁棉1号接菌,鉴定品种对枯萎病菌的抗病性及其与含糖量的关系。试验结果表明,含糖量愈高,感病愈重;含糖量愈低,抗病性愈强,其分界值还原糖含量约为样品干重的12%,水溶性总糖为样品干重的15%(表3-9)。棉花枯萎病是一种高糖病害。
品种含糖量(%)(干重)抗、感枯萎病表现*还原糖水溶性糖病指反应型鲁棉一号15.12117.8535.07S(感)岱字棉1513.24917.2234.62S徐州14212.11015.2129.44S陕115512.01713.739.69R8618.95812.377.92R(抗)
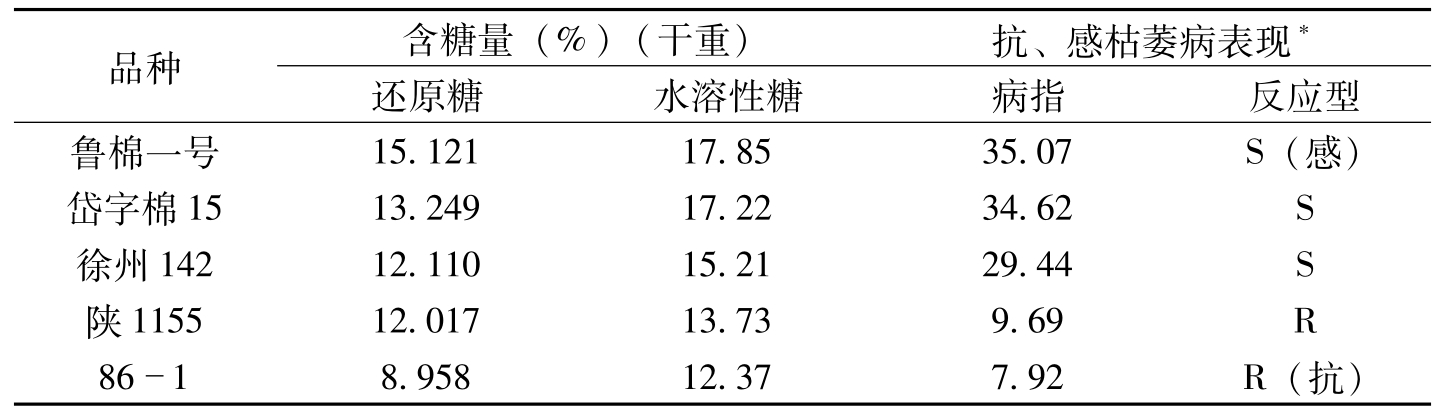
表3-9 不同棉品种苗期含糖水平与抗枯萎病性表现
冯洁等(1991)报道,抗病品种(86-1、陕1155)和感病品种(岱字棉15、徐州142)接菌后,果糖、蔗糖含量均为上升趋势。接菌后抗、感病品种在葡萄糖、核糖含量上存在差异,前者接菌后葡萄糖、核糖含量明显上
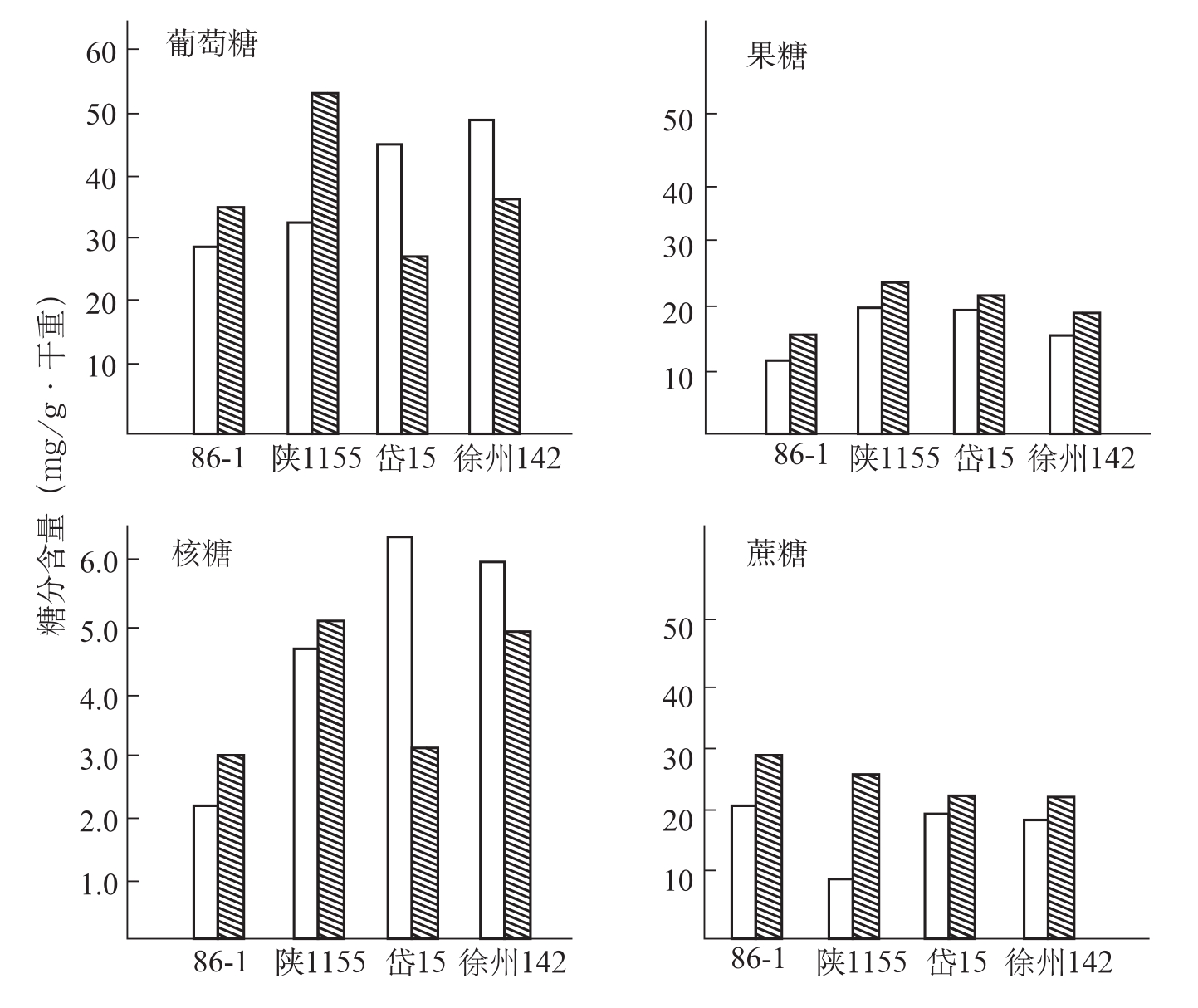
图3-11 棉花叶内糖分含量变化
吴小月等(1993)和宋凤鸣等(1996)的研究结果也表明,感病品种棉苗体内的含糖量高于抗病品种(表3-10,表3-11)。
品种泗棉2号岱红岱盐棉488618311中棉所12子叶0.4330.4040.3870.2970.2620.259蕾期叶片0.7580.7430.5750.6400.6130.515抗病性SSMRRRR
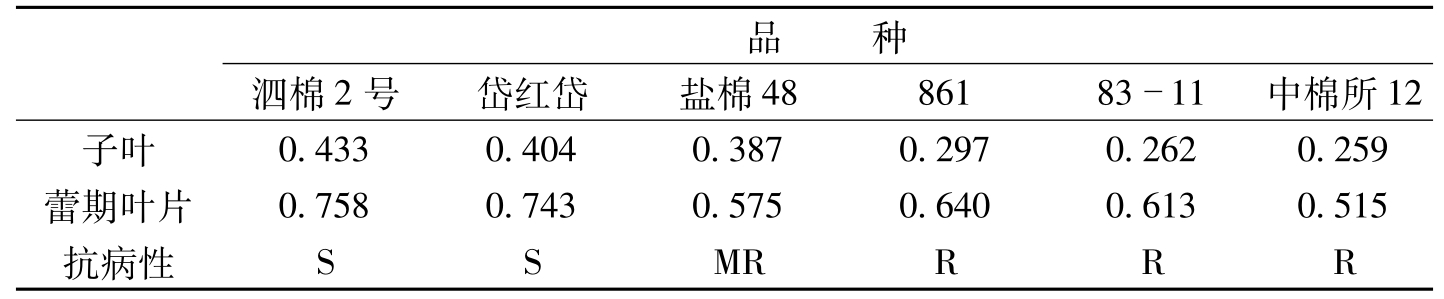
表3-10 不同抗性的陆地棉品种间可溶性糖含量比较(吴小月等,1993)
品种处理叶片中葡萄糖含量(mg/g鲜重)根茎部中葡萄糖含量(mg/g鲜重)总糖可溶性糖还原糖总糖可溶性糖还原糖中棉所12(抗病)不接种17.599.332.577.974.141.98接种25.5213.315.5718.5410.432.646037(感病)不接种16.2010.922.7710.004.452.44接种24.8214.827.1520.7110.344.07
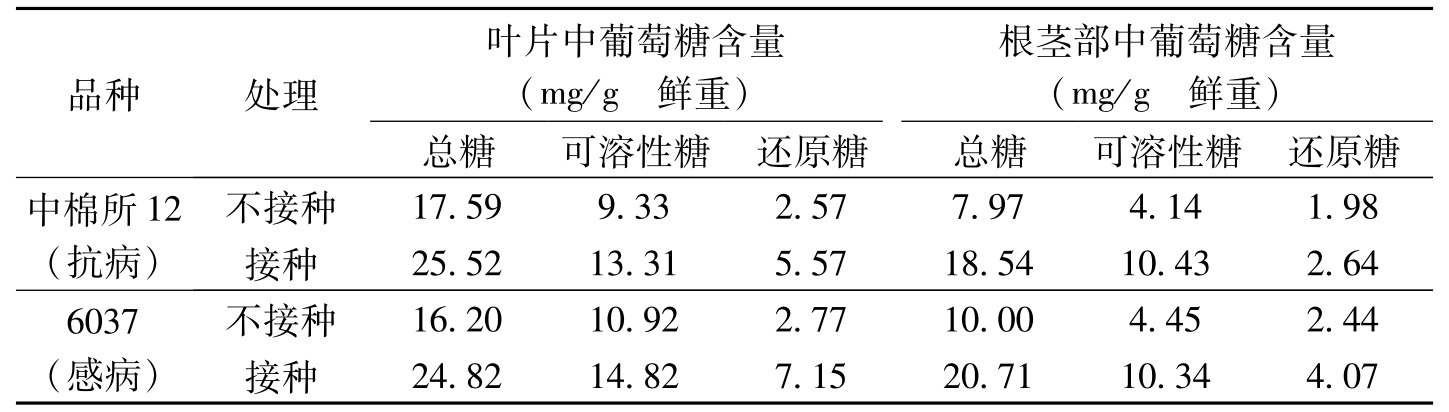
表3-11 棉苗组织中糖含量的变化(宋凤鸣等,1996)
3.酚类
受病原物侵染后,植物体内可溶性酚类物质会大量积累,大量的酚类物质通过延缓入侵病原物的生长而在植物抗病性中起作用。细胞壁上存在大量的酚类物质,这些不溶性胞壁结合酚及细胞壁的快速木质化与植物抗病性有密切关系。棉花体内含有丰富的酚类(主要是多元酚)物质,随着棉苗生长其酚类物质含量增加,同时,可提高对病害的抗性。
多酚类物质在棉花抗病性具有重要的作用。刘发敏等(1993)报道,在不接枯萎病菌的条件下,棉花中多酚类含量是感病品种高于抗病品种,但抗、感病品种的多酚类含量都是随着生育期进程由低到高。抗病品种86-1在1叶期为1.41%,开花期为2.99%;感病品种鲁棉1号在1叶期为3.25%,开花期为4.02%。鲁棉1号在各生育期均高于86-1。但在接枯萎病菌的条件下,86-1多酚类含量几乎是直线上升,1叶期为1.62%,开花期为4.38%,开花期为1叶期的2.7倍。鲁棉1号相反,多酚类含量从1~3叶期下降较大,以后下降减缓。1叶期为3.65%,开花期为2.91%,开花期仅为1叶期的4/5。86-1和鲁棉1号相比,从5叶期开始,86-1高于鲁棉1号,到开花期为鲁棉1号的1.5倍。袁章虎等(1995)指出,各品种无论是抗病的还是感病的,接种枯萎菌后体内多酚的含量都有明显提高。抗、感病品种主要差异有以下两点:一是抗病品种多酚的累积高峰出现的早;二是抗病品种在接菌后多酚增加的灵敏量比感病品种高许多。这表明品种所固有的组成性多酚含量是品种间的差异,在一定范围内与抗病性没有关系,而棉株遭受枯萎萎菌侵染后所合成的多酚类物质才是抗病性增强的根本所在。
病原物侵染后,植物体内常有大量酚类物质的积累,这些酚类物质主要由苯丙烷类代谢途径合成,其中,绿原酸和阿魏酸是主要的酚类物质。作为苯丙烷类代谢途径的主要产物,在受病菌侵染或诱发物处理后,植物体内绿原酸和阿魏酸的积累是一种常见的现象。在一些植物的抗病反应中绿原酸含量增加快,发生早,而感病反应中则相反。在绿原酸方面,冯洁等(1990)试验结果表明,不同抗病类型品种接种后绿原酸含量存在差异。抗病品种中棉所12、86-1在接菌后24h绿原酸含量迅速上升,最高含量0.47~0.51mg/g干重,48h后又迅速下降,以后又有所回升。感病品种岱字棉15、豫棉1号在接菌初期(24~72h)绿原酸含量始终低于未接菌的对照,直到96h才迅速增加,最大含量为0.49~0.55mg/g干重。在接菌初期感病品种体内酚类植保素积累很慢,到后期含量虽然超过了抗病品种,但已错过了杀伤病菌的时机。抗病品种则不然,在接菌初期绿原酸含量就迅速积累,对病菌的侵入起到阻止和杀伤作用。可见接种枯萎菌后,抗、感品种都可以产生酚类植保素——绿原酸,关键在于产生的量和速度不同。宋凤鸣等(1996)的研究结果也表现了这种特点。Friend等(1973)研究表明,绿原酸可抑制某些病原菌的生长和产孢。阿魏酸虽然无直接的杀菌活性,但它是木质素的前体,其含量的增加可为木质素合成提供更多的底物,促进木质素的积累,导致细胞壁的木质化,从而在植物抗病性中起了间接作用。冯洁等(1990)研究结果表明,抗病品种中棉所12、86-1在接菌后48h均出现了一个阿魏酸增加峰,显著高于对照水平,最大量可达4.27~6.68mg/g干重,86-1在接菌后96h又出现一个增加峰,含量可达5.01mg/g干重。而感染病品种岱字棉15、豫棉1号在接菌后24~72h,阿魏酸含量始终低于未接菌的对照,直到96h才略高于对照,最大量为2.17~3.48mg/g干重。接菌后阿魏酸含量的变化与棉花抗枯萎病性呈正相关。宋凤鸣等(1996)报道,在不接种枯萎病菌条件下,抗、感品种棉苗体内阿魏酸含量明显差异,但接菌后,抗病品种棉苗体内阿魏酸含量的增加幅度更大(表3-12)。
处理中棉所12(抗病品种)6037(感病品种)接种后7天接种后11天接种后15天接种后11天接种后15天不接枯萎病菌0.5820.5450.7520.5720.692接枯萎病菌1.3021.7592.1130.8031.201
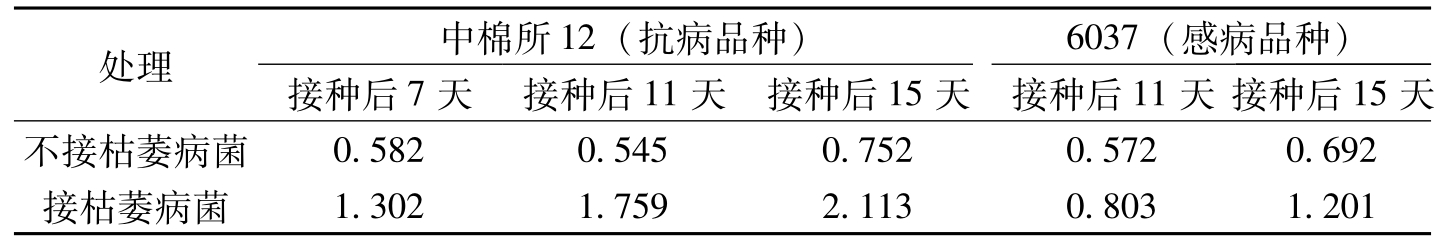
表3-12 棉苗根茎部组织中阿魏酸的含量(OD350/gDW-ml)
由于病原真菌和细菌感染,在受损害的植物细胞内或细胞周围高浓度地积累植保素,这些小分子化合物许多是黄酮类化合物。黄酮类化合物以糖苷的形式广泛分布在植物界。它们的合成和转化可能彼此独立地加以调节。刘发敏等(1993)指出,在不接枯萎病菌的条件下,抗、感病品种黄酮类含量随着生育期进程变化较大。抗病品种86-1每克干重在一叶期为10.5473mg,三叶期是高峰为14.0084mg,开花期为9.3064mg;感病品种鲁棉1号每克干重在一叶期为9.3272mg,现蕾期为高峰是12.5100mg,开花期为11.6401mg,86-1在1~3叶期高于鲁棉1号,其余各期都低于鲁棉1号。在接菌的条件下,86-1黄酮类含量每克干重在一叶期为10.7138mg,到五叶期降到最低点为7.2778mg,然后急剧上升,开花期为12.2706mg,曲线呈现“V”字形。鲁棉1号黄酮类含量每克干重在一叶期为9.8407mg,三叶期达到高峰为11.5603mg,现蕾期为10.2887mg,开花期为10.446mg,变化较平缓,曲线略呈“S”形。86-1和鲁棉1号相比,在三叶期和五叶期86-1黄酮类含量低于鲁棉1号外,其余各生育期都高于鲁棉1号(图3-12)。宋凤鸣等(1996)也报道了类似的研究结果。接种枯萎病菌的抗病品种棉苗叶片和根茎部组织中黄酮类物质含量略高于感病品种,枯萎病病菌侵染后,棉苗组织中黄酮类物质含量明显提高,抗病品种棉苗组织中黄酮类物质含量的增加幅度略大(表3-13)。
品种处理叶片组织根茎部组织中棉所12号(抗病)不接枯萎病2.041.212.12接枯萎病3.622.643.546037(感病)不接枯萎病1.691.091.68接枯萎病3.031.802.70
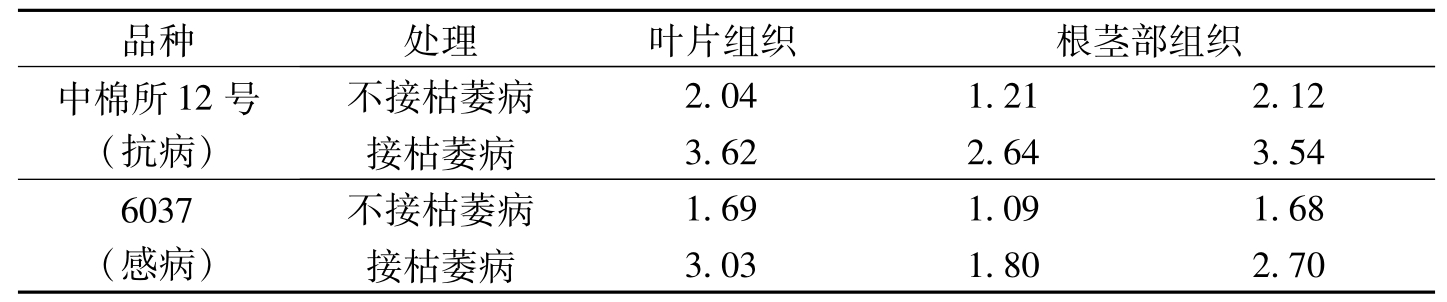
表3-13 棉苗组织中黄酮类物质的含量(OD509/g·ml min)
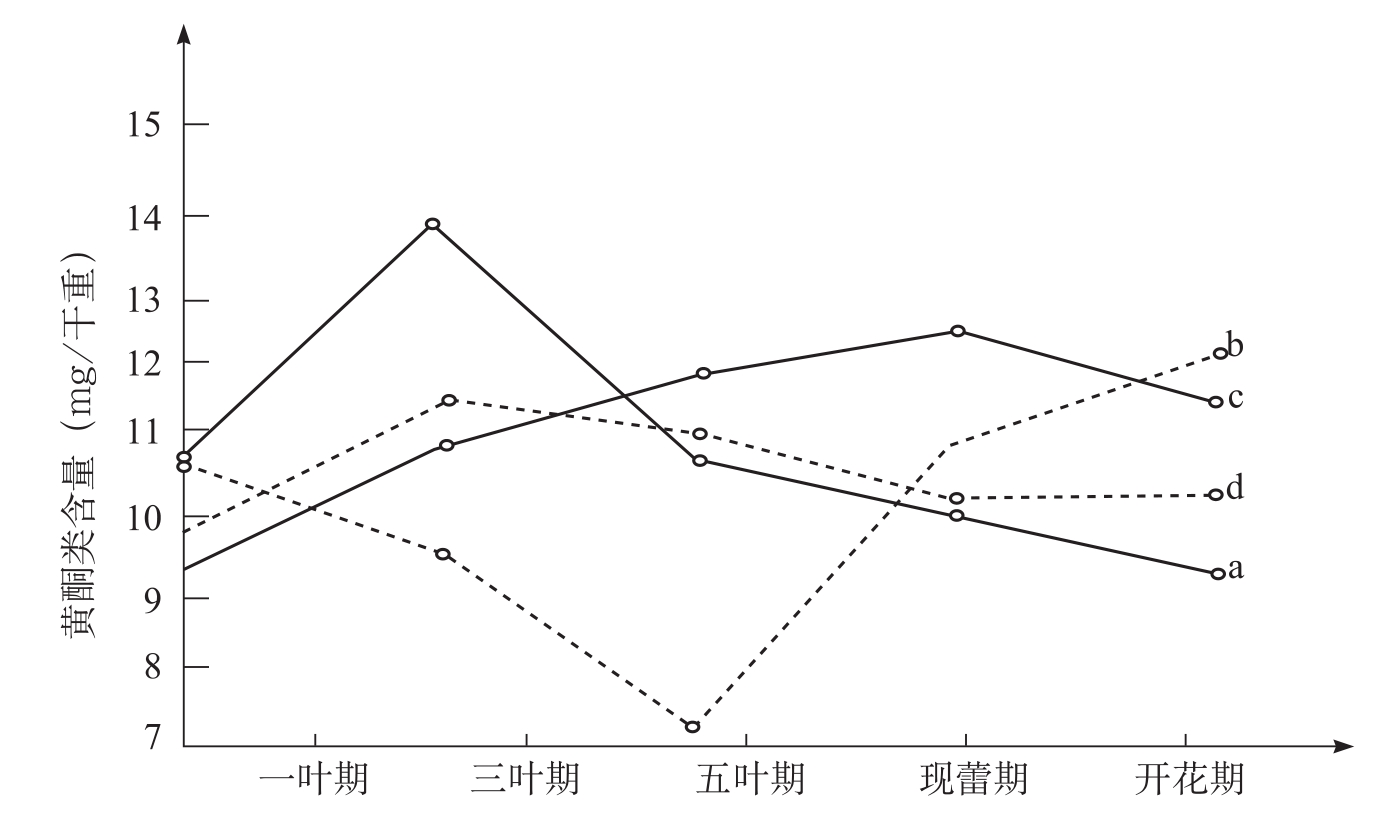
图3-12 棉花不同生育时期黄酮类含量的变化
植物细胞壁上存在着众多的酚类物质。在一些植物病害系统中已经证实植物受侵后组织中不溶性胞壁结合酚类物质在其对抗病性中起重要作用。Glazener等(1982)认为不溶性胞壁结合的简单酚类物质可能参与植物的某些防卫反应过程,且与抗病性有关。Nieman等(1991)和Bonello等(1993)报道,胞壁结合的复杂酚聚合物主要是木质素,其含量的增加,表明细胞壁的木质化,因而在植物抗病性中起到作用。宋凤鸣等(1997,2001)研究结果表明:①抗病品种棉苗组织中胞壁结合的简单酚、酚聚合物及黄酮醇含量高于感病品种棉苗组织中的含量。根茎部组织中胞壁结合酚含量相对较高;②枯萎病菌侵染后棉苗组织中不溶性胞壁结合酚含量有明显的提高(表3-14)。因此认为,受枯萎病菌侵染后棉苗组织中可溶性酚及不溶性胞壁结合酚的积累与棉苗对枯萎病的抗性有关。
品种处理叶片组织根茎部组织简单酚复杂酚聚合物黄酮醇简单酚复杂酚聚合物黄酮醇中棉所12号(抗病)不接枯萎病菌0.1900.1132.890.3150.2384.72接枯萎病菌0.3640.1964.810.5610.4357.506037(感病)不接枯萎病菌0.1570.0882.610.2990.1984.20接枯萎病菌0.2440.1443.530.4290.3386.31

表3-14 棉苗接种后11天时单位干细胞壁组织[m(DW)]中不溶性胞壁结合酚类物质的含量
儿茶素(Catechin)是棉花植株中最主要的多元酚(Howell等1976),随着棉苗生长,组织中儿茶素的含量增加,同时也提高对立枯病的抗性(Hunter等,1974)。在离体条件下,儿茶素可抑制立枯病菌(
品种处理叶片组织根茎部组织中棉所12号(抗病)不接枯萎病菌0.5170.582接枯萎病菌1.4521.172不接枯萎病菌0.6790.706接枯萎病菌1.7681.6056037(感病)不接枯萎病菌0.4050.528接枯萎病菌0.8520.771
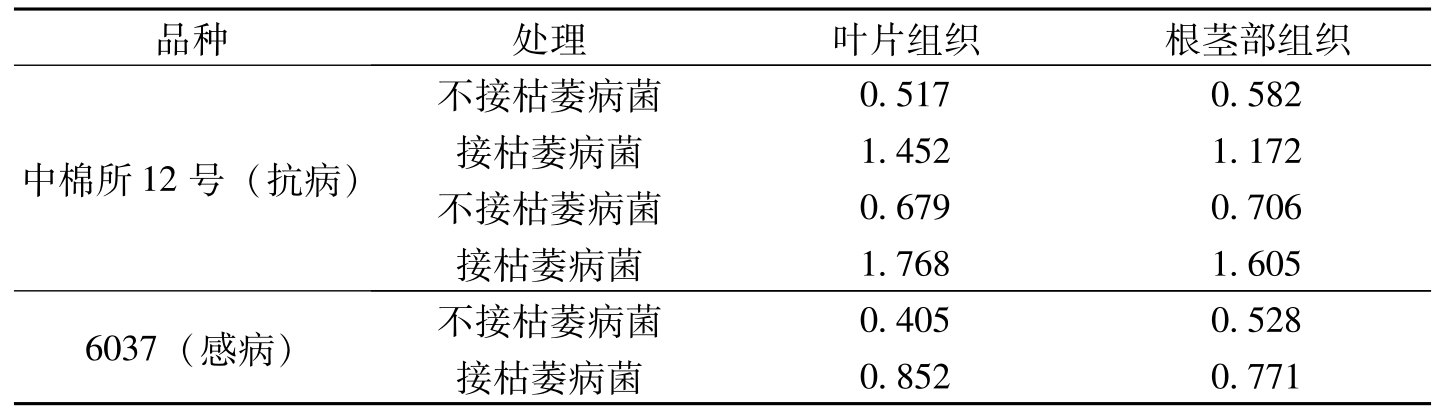
表3-15 棉苗组织中儿茶素的含量(mg/g 鲜重)
儿茶素浓度(mg/ml)菌落直径(cm)菌丝干重(mg)C+S(1)C+NaPP(2)C+SC+NaPP产孢量(%)(×107/ml)孢子萌发(%)0.14.073.47307220757.995.350.053.703.17273199646.790.380.103.773.37257197584.477.680.503.432.67210177553.267.371.002.371.3711384130.551.802.001.120.7797611.433.58EC501.341.121.151.180.791.36
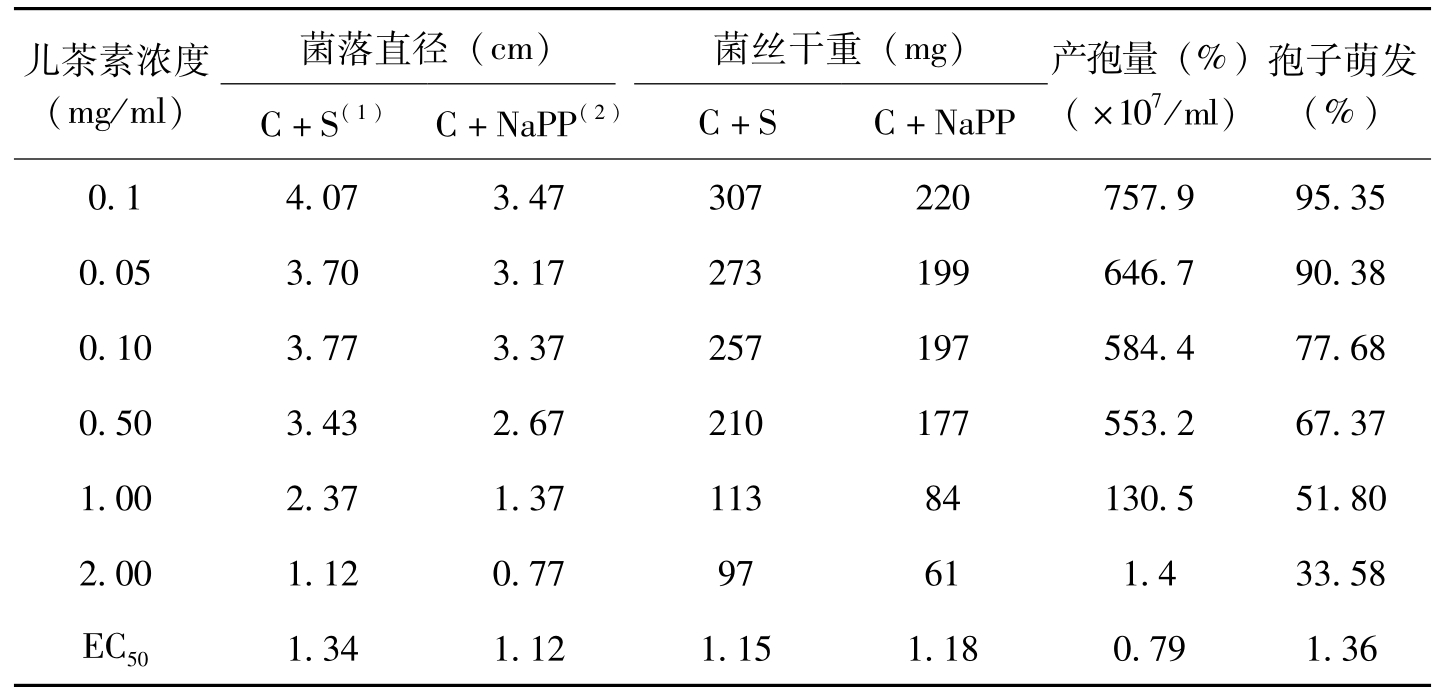
表3-16 离体条件下儿茶素对棉花枯萎病菌的影响
品种处理对PG的抑制作用*(%)对PL的抑制作用(%)叶片组织根茎部组织叶片组织根茎部组织中棉所12不接枯萎病菌30.3423.0819.5518.91接枯萎病菌54.1855.3451.7548.62不接枯萎病菌24.3819.2016.0712.77接枯萎病菌42.7645.3835.4543.57

表3-17 棉苗组织提取液中酚类物质对多聚半乳糖醛酸酶和果胶裂解酶活性的抑制作用
处理PG活性PG(μgglucose/mgprot·h)PL活性PL(△OD550/mgprot·h)对照CK50%926.3091.0含儿茶素部分抽提液673.00(27.32%)*40.3(55.71%)其余部分抽提液874.00(5.62%)86.7(4.76%)
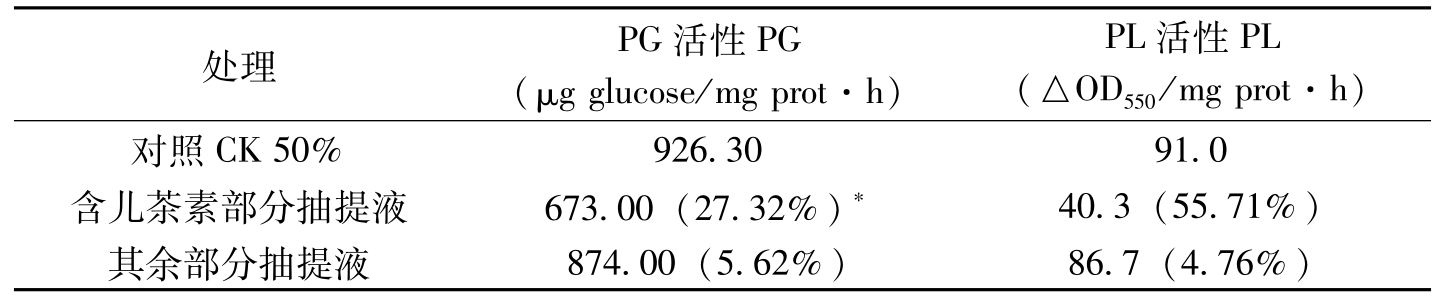
表3-18 组织提取液经TLC后含儿茶素部分硅胶抽提液及其余部分硅胶抽提液对多聚半乳糖醛酸酶和果胶裂解酶活性的影响
棉酚是棉花体内主要的酚类物质,在离体条件下,棉酚可抑制黄萎病菌和枯萎病菌某些专化型的菌丝生长和产孢(Bell,1967;Davis,1964),棉花受病原菌侵染后,其体内棉酚含量有明显的增加,因而推测,棉酚及单宁在棉花抗病性中起到作用(Bell,1967;魏守军等,1992;沈其益等,1992;Harrison等,1982)。魏守军等(1992)观察到在棉花受到枯萎菌侵染后,棉酚在棉株木质部和叶柄维管束导管细胞内大量累积(图3-13),并且抗病品种中棉所12和86-1中,染病后根系游离棉酚含量分别提高112%和96%;而感病品种冀棉11号和鄂荆92中,染病后根系游离棉酚含量只分别提高8%和4.8%。宋凤鸣等(1997)也观察到在未接种枯萎病菌的健康棉苗中,抗病品种的游离棉酚含量均显著高于感病品种相应组织中的含量,根茎组织内的含量又高于叶片;接种棉枯萎病菌引起发病并导致棉苗组织中游离棉酚含量明显提高,抗病品种增长显著高于感病品种。
图3-13 棉株茎与叶柄横切片图
4.蛋白质类
棉花枯萎病是重要的土传病害,枯萎菌由根系直接侵入,根系细胞壁是它需要克服的第一道防线。蛋白质是病原菌赖以生存的重要营养来源,寄主植物细胞壁蛋白质含量十分丰富,一些蛋白质组分(如天冬氨酸、谷氨酸等)经过脱氨后的碳骨架可以直接进入病原菌的代谢系统,合成病菌所需要的蛋白质,因此,对寄主细胞内蛋白质组成成分的氨基酸进行研究的重要意义是显而易见的。冯洁等(1991,1994,1995)在这方面做了较系统的研究,取得了具有科学意义的研究结果。
(1)棉花根、叶内的蛋白质含量
在感染枯萎病菌后,叶内蛋白质含量,抗病品种接菌后36h以前的蛋白质含量都低于未接菌的对照,36h后则高于对照的水平,表现为先低后高;而感病品种则恰恰相反,出现先高后低的现象。在根内,抗病品种接菌后24h始终保持低于对照品种中蛋白质的水平,直到36h后才有所上升。而感病品种接菌后蛋白质浓度均高于对照。另外,达到高峰时的蛋白质含量感病品种高于抗病品种。
(2)棉花细胞壁氨基酸含量
用不同致病力的棉花枯萎病菌7号和3号小种接种棉花,对抗、感品种根部细胞壁氨基酸含量进行分析。结果表明,感病品种在接种7号小种后,根内细胞壁氨基酸含量增加了32.3%,抗病品种增加了4.4%;接种3号小种后抗病品种氨基酸含量下降了14%,感病品种上升了19.8%(图3-14)。感病品种受枯萎菌诱导后细胞壁氨基酸的积累明显高于抗病品种,为枯萎病菌的生长提供了较多的氮源,易与枯萎菌建立寄生关系,这表明细胞壁氨基酸含量与棉花对枯萎病的抗性具有一定的相关性。
(3)棉花细胞壁富含羟脯氨基酸糖蛋白含量
脯氨酸是一种非常重要的氨基酸,它经过羟化形成的羟基脯氨酸,是细胞壁伸展蛋白的原料。有些研究结果表明,细胞壁富含羟脯氨酸糖蛋白的积累与植物抗病性关系密切(Mayad,1986;Benhamou,1990)。冯洁等(1995)研究结果表明,不同品种间细胞壁富含羟脯氨酸糖蛋白(HRGP)的积累量存在差异,抗病品种细胞壁内HRGP含量明显高于感病品种。棉花叶内的HYP(羟脯氨酸)含量与其他氨基酸相比含量最低,只占总量的
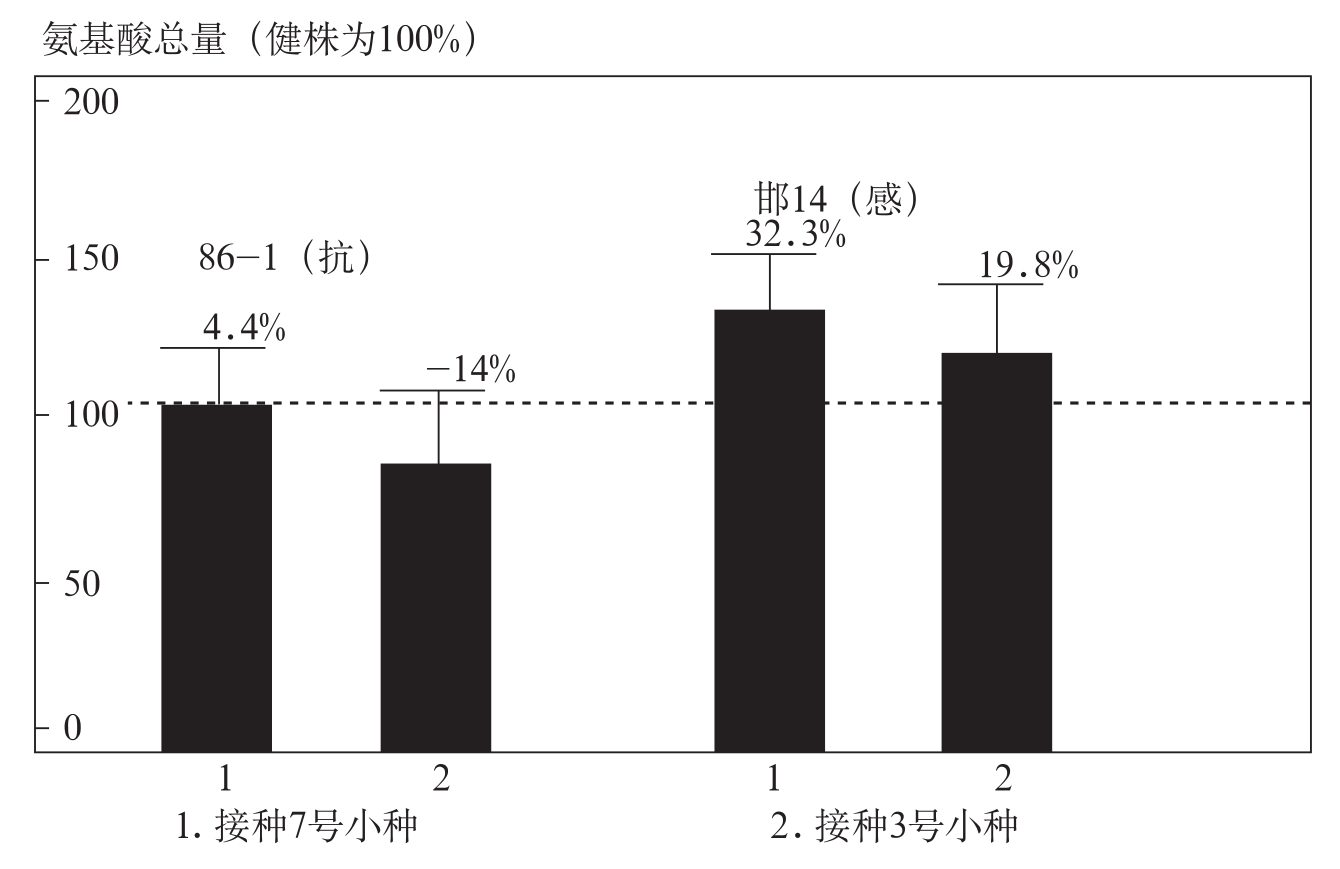
图3-14 接种枯萎病菌对棉花根内细胞壁氨基酸含量的影响
处理861(抗病)邯14(感病)根叶根叶木质素增长率(%)木质素增长率(%)木质素增长率(%)木质素增长率(%)健株2.882.402.802.477号小种3.5623.62.6811.73.2415.72.673.13号小种3.2613.22.566.73.079.62.553.2
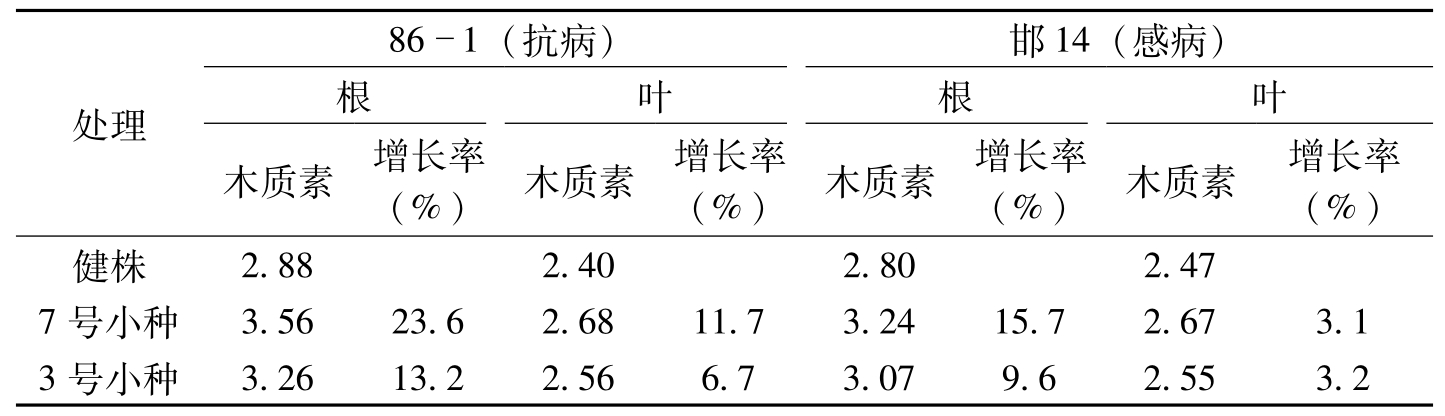
表3-19 不同处理对细胞壁木质素沉积的影响
至于HRGP在抗病反应中的作用机制主有以下3方面(宋凤鸣,1992)。
(1)作为凝集素的作用
许多植物凝集素是糖蛋白,其中,有一些就是HRGP。这种具凝集活性的HRGP定位于能与病原菌相互作用的位点,HRGP可能与病原菌相互作用并把病原菌固定在细胞壁中,从而阻止病原菌的侵入或在细胞间的扩散。HRGP还在寄主—病原菌相互作用的专化性识别机制中起到重要作用。
(2)作为木质素的沉积位点
寄主与病原菌相互作用中,细胞壁在病原菌侵染后的木质化是寄主抗病反应的特性之一。木质素的形成与苯丙氨酸代谢形成的松柏醇有关,松柏醇的脱氢多聚物(DHP)与HRGP结合形成一种对酸稳定的复合物。在初生壁的木质化过程中,由松柏醇产生的甲基化醌与HRGP之间形成共价交错连接,从而导致了木质素在细胞壁中的沉积。
(3)作为结构屏障的作用
高等植物细胞壁中,HRGP填充在纤维素骨架的间隙中,又与细胞壁的其他成分共价结合形成更为致密、不可穿透的结构屏障。而且,HRGP具有结构性多聚物的功能,从而提高了细胞壁的强度。在侵染穿透过程中,病原物分泌的纤维素酶、半纤维素酶和蛋白酶等都能分解纤维素、半纤维素等细胞壁成分。但这些酶不能分辨HRGP,而且HRGP包围在纤维素和半纤维素的周围,从而把病原物分泌的酶与其底物分开,使纤维素和半纤维素等胞壁物质免受分解。继续保持细胞的正常结构,阻止病原物的侵入。但目前尚没有这方面的直接证据。
5.活性氧及膜脂过氧化
病原菌的侵染可破坏寄主植物体内活性氧产生与清除之间的动态平衡,引起活性氧(主要是超氧阴离子O
活性氧及膜脂过氧化的产物在植物抗病防卫反应中起的主要作用是:①对入侵病菌的抑制;②诱导植保素合成、细胞壁木质化及富含羟脯氨酸糖蛋白的沉积;③与信号传递有关并激活植物防卫反应相关基因的表达。宋凤鸣等(2001)研究结果表明,未接种枯萎病菌的健康棉苗中,抗病品种中棉所12棉苗的活性氧水平略高于感病品种6037,接种病菌后,棉苗叶片和根茎组织的活性氧水平显著升高,中棉所12的活性氧积累早,积累水平高于6037(表3-20)。健康棉苗的茎与根组织中LOX酶活性,在抗与感病品种间无明显差异,接种枯萎病菌后,2个供试品种棉苗组织中LOX酶活性有明显的升高,但表现不同的变化动态。抗病品种中棉所12接种后茎与根组织中,LOX酶活性快速增加,在接种后5~8天达最高,之后缓慢下降。感病品种6037棉苗接种后茎与根组织中,LOX酶活性增加缓慢,8天后才有较大幅度的增加,而后逐步下降。
处理O-2产生速率(μmolO-2/t(gFW·min)H2O2积累(μmolH2O2/gFW)3天7天3天7天中棉所12(抗病)不接枯萎病菌0.320±0.0050.381±0.037103.9±4.486.3±8.8接枯萎病菌0.922±0.0351.086±0.066163.0±3.5159.6±4.56037(感病)不接枯萎病菌0188±0.0010.253±0.00661.5±7.852.9±4.6接枯萎病菌0.336±0.0180.725±0.00685.6±3.1127.5±5.3
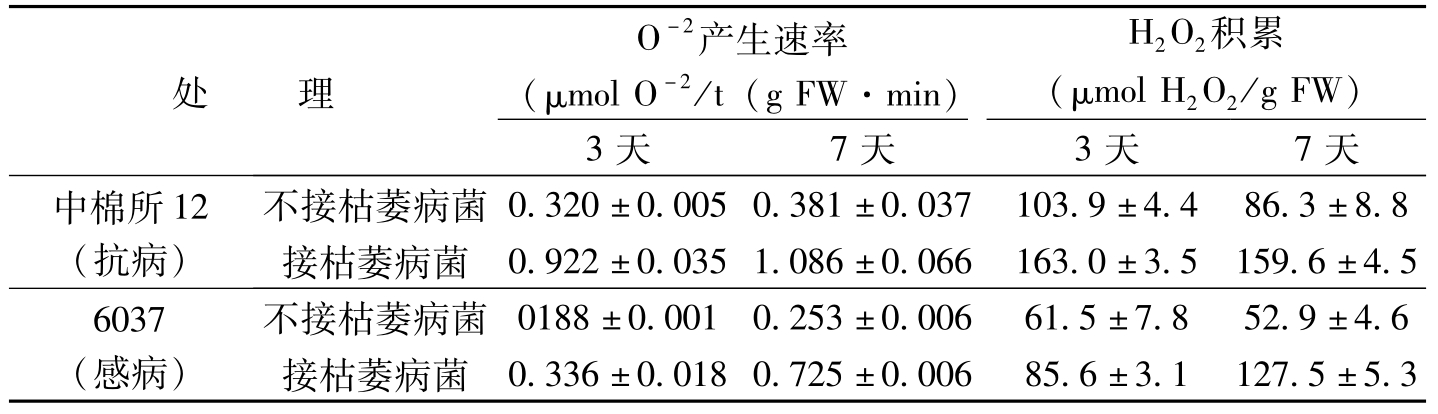
表3-20 枯萎病菌接种后棉苗茎与根组织中活性氧的积累
丙二醛(MDA)是膜脂过氧化产物,未接种枯萎病菌的健康棉苗的茎与根组织中,MDA含量在抗与感病品种间无显著差异,但枯萎病菌接种后,棉苗组织中MDA的含量有明显的增加,抗病品种中棉所12棉苗MDA的含量在接种后3天就明显增加,此后直线上升,而感病品种6037棉苗中MDA的含量在接种后7才开始积累,增加速度相对较慢。接种枯萎病菌后,抗病品种棉株体内膜系统中,不饱和脂肪酸含量下降明显,而感病品种无显著变化。膜脂过氧化产物丙二醛(MDA)含量上升,膜系统中,不饱和脂肪酸含量下降,表明有膜脂过氧化的发生(宋凤鸣等,2001)。
上述这些结果说明,枯萎病菌侵染后棉苗体内活性氧的积累、LOX酶活性的上升以及由此引起的腆脂过氧化,可能在棉苗对枯萎病的抗性中起
6.生理代谢
棉株感枯萎病后显示的病症是由于一系列不正常的生化和生理活动的结果。脂肪酸是细胞质膜的主要组织成分,具有极重要的生物功能,棉花叶片中的脂肪酸主要由两种饱和脂肪酸和3种不饱和脂肪酸所组成。感染枯萎病棉株叶片中亚麻酸含量下降,而油酸和亚油酸含量增加,总的来看是不饱和成分降低,这说明叶片膜脂中脂肪酸的组成分发生了变化、加之其他生理代谢的影响,导致病株呈现皱缩(宋晓轩等,1992)。郭金城等(1990)测定结果指出,子叶中油酸和亚麻酸含量与品种的抗枯萎病性有一定的相关性。品种的抗病性越强,其子叶中油酸含量越高。高抗枯萎病品种52-128子叶中油酸含量比感病品种鄂沙28高16.99%,表明品种的抗病性与子叶中油酸含量呈正相关;而子叶中亚麻酸含量与品种抗病性呈负相关趋势,子叶中亚麻酸含量越低,其品种的抗病性越强。高抗枯萎病品种52-128子叶中亚麻酸的含量比感病鄂沙28低10.55%。
枯萎病对棉株光合作用和呼吸作用有明显的影响。丁钟荣等(1988)测定结果表明,无论感病品种还是抗病品种,感病植株的光合作用均显著降低。感病品种的光合强度降低了37.2%,总光合强度降低30.5%;抗病品种的光合强度降低28.5%,总光合强度降低21.3%。说明感病植株生长矮化和产量降低都与光合下降有关。净光合速率表明叶片吸收固定二氧化碳的能力,受叶龄影响较大,反映在主茎叶上即是受不同叶位的影响。健株叶片的净光合速率最大值均出现在第五叶位叶片,而病株叶片则无此规律,这是由于其各叶位叶片发病程度和发病时间的不同所致。相同叶位的主茎叶片的净光合速率,病株均明显低于健株。病株叶片的净光合速率平均比健叶下降了58.7%~62.8%。这表明,病叶的同化二氧化碳能力大为降低。
棉株感病后呼吸强度均明显下降,抗病品种下降更甚。抗病品种的病株呼吸强度比健株下降23.7%,而感病品种的病株只比健株下降13.1%。气孔是二氧化碳和水气进出叶片的通道,气孔导度表明气体通过气孔传导的能力,直接影响着气体交换的进程。棉花感染了枯萎病后,各主茎叶位叶片的气孔导度明显降低,病叶的气孔导度分别比健叶的平均降低了
温度是维持正常新陈代谢的重要因素之一。宋晓轩等(1992)测定结果表明,感染枯萎病后,各节位主茎病叶叶片的叶温均高于健叶,最多能高出5.5℃;健叶叶温比其周围环境的气温低2.51~3.13℃,叶温与气温温差较大,而病叶的叶温与气温温差较小,最多相差-1.29~1.24℃,负值说明叶温超过了气温,最高可超出2℃。这种差异与棉株发病程度有直接关系,对病叶外表健康的部分和发病部分的测量,坏死组织部分的温度要比绿色组织部分的温度高。在幼茎和幼铃上也表现了同一规律。
在棉株体内糖、氮代谢方面,病株的硝态氮含量显著减少。感病品种硝态氮含量比健株减少1/2;抗病品种减少1/3。感病株可溶性蛋白质含量显著降低(图3-15)。可溶性糖含量感病植株比健株减少一半左右。同样,抗病品种比感病品种高出1/3以上(丁钟荣等,1988)。张海娜等(2011)报道,感病株体内可溶性糖含量高于正常株(图3-16)。表明抗病品种的营养积累要比感病品种优越得多。
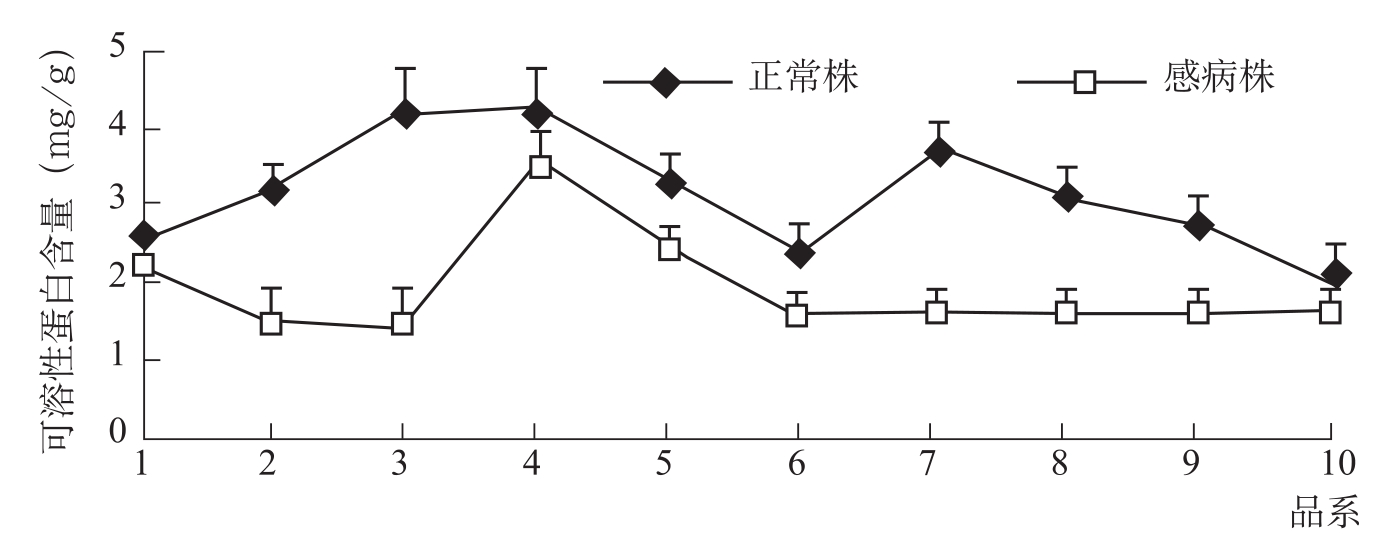
图3-15 可溶性蛋白质含量的变化(张海娜等,2011)

图3-16 可溶性糖含量的变化(张海娜等,2011)
(三)生态抗性
已有的研究结果表明,根际微生物和棉际分泌物等生态因素与植物抗病性有密切关系。植物的分泌物主要包括糖、氨基酸、蛋白质、维生素、有机酸、无机离子等。这些分泌物中的某些物质能在抵御病菌侵入中起作用。显然植物的分泌物中存在着某些物质,由于它们的存在使植物体本身具有抗病的潜在活性,这是植物体自身防御作用的机理之一。
冯洁等(1991)研究了棉花抗、感品种的病、健株根分泌物中氨基酸及糖分含量的变化及其对枯萎菌孢子萌发的影响。结果表明,抗病品种健株根分泌物中,含有7~8种氨基酸,总量分别为1.40nmol和1.33nmol。感病品种根分泌物中,含有1~4种氨基酸,总量分别为0.43nmol和1.02nmol。接种枯萎菌后,棉花抗、感品种根分泌物中,氨基酸种类均达到17种,抗病品种根分泌物中,氨基酸总量为152.40~160.30nmol。感病品种为216.10~239.98nmol。感病品种根分泌物中氨基酸总量明显高于抗病品种。丙氨酸(Ala)在氨基酸总量中所占的比例最大,其次是谷氨酸 (Glu)和缬氨酸(Val),三者之和在氨基酸总量中所占的比例抗病品种为60%~75%,感病品种为59%~64%。天冬氨酸(Asp)和谷氨酸(Glu)是植物及病原物体内代谢的重要氮化物。感病品种接菌后根分泌物中,这
孢子在不同氨基酸溶液中萌发,在Gly.Ala、Ser、Ghl中,孢子萌发百分率最高(61.0%~74.5%),其次是Asp、Phe,Thr、Pro、Val、Cys萌发率在30.2%~58.4%,也具有刺激孢子萌发的作用,在Arg、Met、Leu中,萌发率接近清水对照(22.6%~27.7%),只有His具有抑制孢子萌发的作用(17.2%)。
棉苗健株根分泌物中,总糖含量很低,用薄层层析法检测不到果糖和葡萄糖。接菌后总糖含量与健株相比有大幅度增加,并可检测到果糖和葡萄糖。抗病品种的果糖含量分别为23.70%~25.7%和2.69%~3.02%;而感病品种果糖和葡萄糖的含量分别为25.20%~25.60%和1.82%~1.92%。抗病品种的葡萄糖含量略高于感病品种(冯洁等,1991)。
由于棉花根分泌物的成分非常复杂,其作用在土壤中又受其他诸多因素的影响,进一步研究这一问题,对探明枯萎病生态抗性机制是必要的。
棉枯萎病菌主要存活于土壤并从根部侵入,根际则是病菌侵入根系的必然通道。而根际微生物是一个极其庞大的群体,其区系各成员对病菌的反应也各不相同,有些表现为抑制作用(包括营养竞争、拮抗、寄生、捕食等),有些表现为促进作用(包括促进萌发,促进生长,协助侵入等),有些则表现关系不大,根际微生物区系的组成不同,对病菌总的反应也就不同。因此,弄清根际微生物的区系组成(包括种类、种数、优势种等),对于抗病机制的研究及生物防治都具有十分重要的理论意义,对于开发土壤微生物资源也有较为重要的作用。李洪连等(1990、1991、1992)对棉花抗、感枯萎病品种根际微生物种类、数量及其抑菌作用进了研究。研究
播期品种生育期真菌平均值及差异比较放线菌平均值及差异比较细菌平均值及差异比较春播861(抗病)冀棉7号(抗病)河南69(感病)鲁棉1号(感病)2~3叶期11.886~7叶期28.34现蕾期13.152~3叶期12.566~7叶期23.95现蕾期13.002~3叶期6.036~7叶期6.99现蕾期6.362~3叶期7.496~7叶期5.36现蕾期5.6017.79aA16.50aA6.46bB6.15bB7.6323.3228.846.2813.4412.444.4913.3010.873.7910.367.9619.93aA10.72bB9.55bB7.37bB13.2719.39102.134.2312.8092.022.986.4656.936.0514.2759.2744.93aA36.36abAB22.12cB26.53bcB
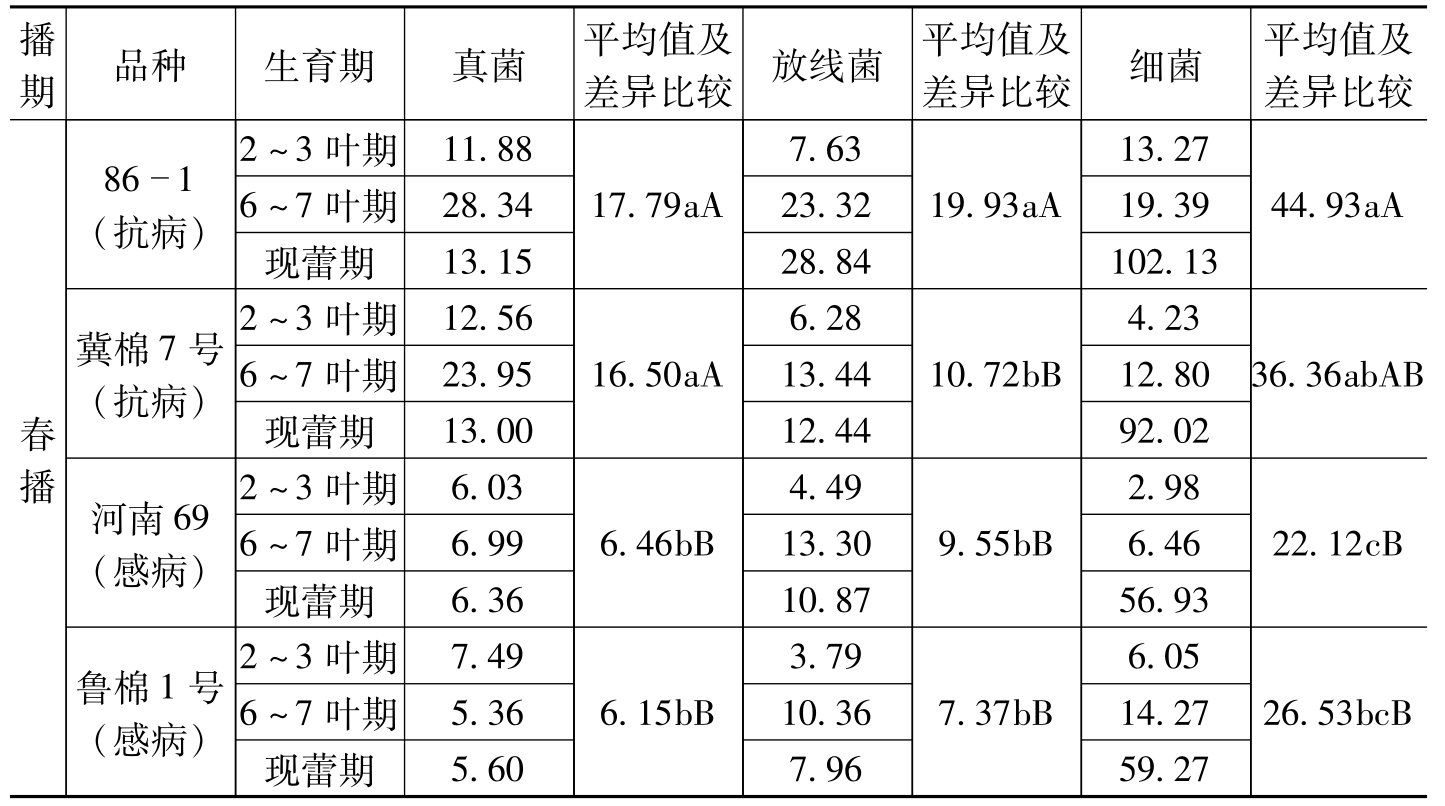
表3-21 棉花抗、感枯萎病品种根际微生物数量(106个/克土)
播期品种生育期真菌平均值及差异比较放线菌平均值及差异比较细菌平均值及差异比较夏播861(抗病)冀棉7号(抗病)河南69(感病)鲁棉1号(感病)2~3叶期22.556~7叶期27.11现蕾期36.392~3叶期18.376~7叶期29.07现蕾期28.172~3叶期10.146~7叶期15.88现蕾期19.842~3叶期8.666~7叶期9.44现蕾期13.7328.68aA25.20aA15.59bB10.61bB13.6914.7016.508.7012.8912.034.146.065.624.944.624.4714.96aA11.21bB5.27cC4.68cC21.7478.45141.658.4751.5998.0612.1537.8590.936.4254.3993.0780.61aA52.71bB46.98bB51.29bB
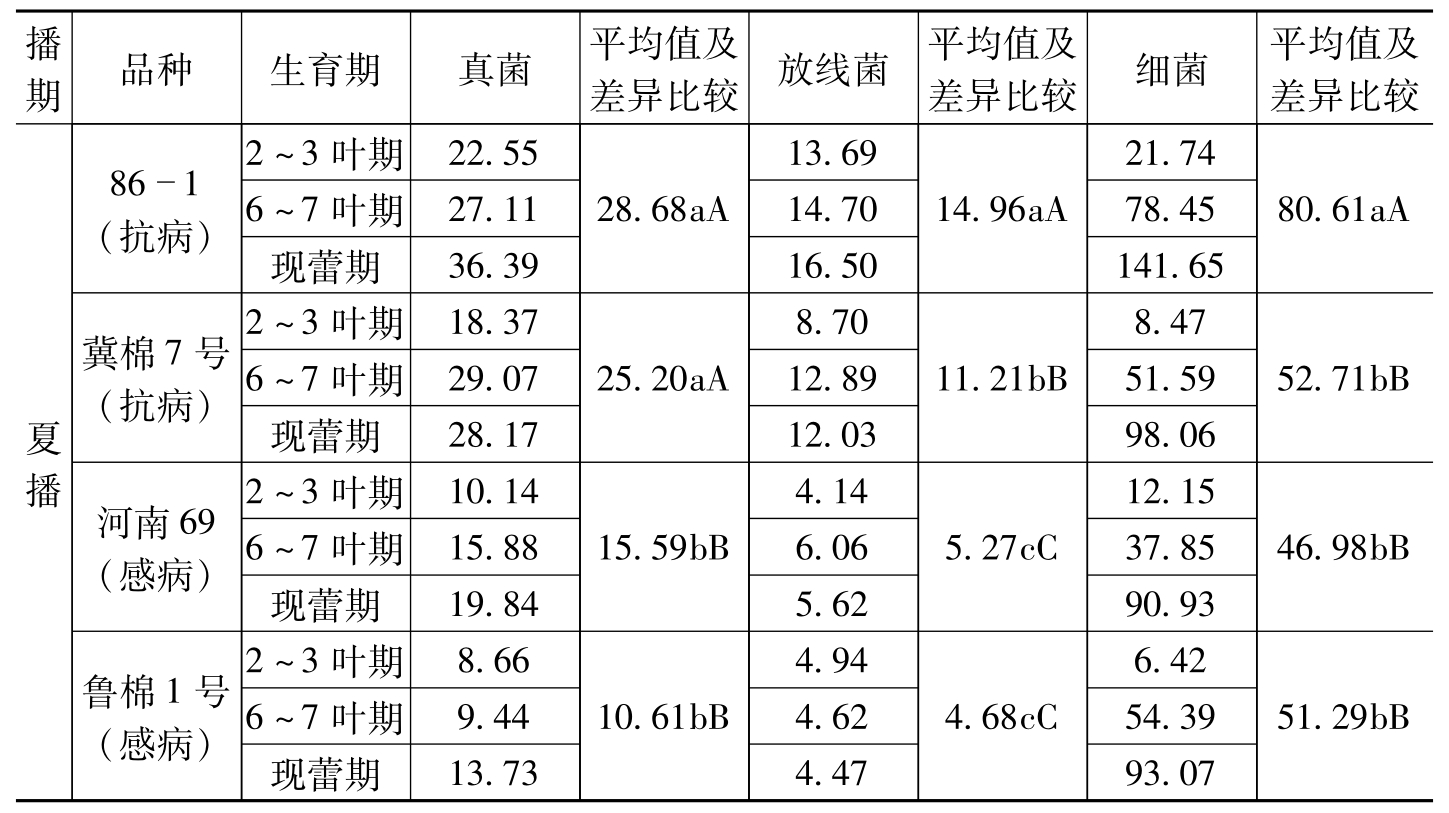
表3-21 棉花抗、感枯萎病品种根际微生物数量(106个/克土)(续)-1
棉花抗、感枯萎病品种在相同条件下根际微生物种类和数量明显不同,其原因可能是由抗、感品种根系分泌物和脱落物的不同而导致的。如果根际内微生物数量较多,就会对枯萎病菌的侵入产生较大的抑制作用,使病菌难以侵入,从而使棉株表现为抗病。当然,这种影响除决定于根际微生物的数量外,还与微生物的种类及对枯萎病菌的抑菌作用强弱有关。
综上所述,棉花枯萎病的抗病机制是一个非常复杂的问题,涉及的因素众多。如果强调某一方面的作用,难免有一定的局限性。抗病、耐病和感病品种不仅在受到病原物侵入前就存在差异,而且更重要的是染病后抗性与其产生抗性机制的速度和程度的差异。如果将有关联的作用有机地联系起来,开展综合研究,可能更有助于深入地认识棉花对枯萎病的抗病机制。









