棉花抗枯萎病育种
棉花抗枯萎病育种
生产实践业已证明,应用抗病品种是枯萎病综合防治技术体系中的核心技术,也是防治枯萎病最经济、有效的技术途径。据1984年的全国棉花枯、黄萎病综合防治研究协作组的统计,全国种植棉花面积约8800余万亩,而黄、枯萎病的发病面积近2200万亩,占调查面积的31.3%,年损失皮棉75万~100万t,折合人民币2亿~3亿元。可是推广了以种植抗病品种为中心的综合防治技术以后,枯萎病的发病率压低到5%以下,挽回了皮棉损失。同时,由于节省了农药等开支,极大地降低防治费用,杜绝了人畜中毒事故和环境污染,保护了棉田生态环境,取得了显著的经济、生态、社会效益。而抗枯萎病育种的成效取决于对病原菌与寄主(棉花)的互作关系和枯萎病抗性遗传规律的认识,抗源的获取以及科学的育种方法。
一、病原菌与寄主(棉花)的交互作用
了解病原菌与寄主(棉花)之间的交互作用,是选育抗棉花枯萎病品种的重要理论依据之一。这里包括两方面内容:一是垂直抗性与水平抗性;二是基因对基因假说。
(一)垂直抗性与水平抗性
这一理论观点由Van Der Plank(1963)提出。
1.垂直抗病性(Verntical resistance)
寄主对某些病原生理小种具有免疫或高抗性,可是对另一些生理小种
2.水平抗病性(Horizontal resistance)
亦可称为非专化抗病性(non-specific resistance)。水平抗性指寄主品种对各个病原生理小种的抗病反应,大体上接近于同一水平,它对病原菌的不同小种没有“特异”反应或“专化”反应,几乎在同一水平线上。它的作用主要表现在能阻止病原菌侵入寄主后的进一步扩展和定植,表现为潜育期较长,病斑小而少,病原菌繁殖体的数量相对较少。因而病害发展的速度较缓慢,程度较轻。在农业生产中,人们长期利用水平抗性,但因其抗、感症状表现不如垂直抗性明显,鉴别较难,过去在抗病育种中往往忽视了具有水平抗性的品种。由于它对病原菌生理小种不形成定向选择的压力,因而不致引起生理小种的变化,也不会导致品种抗性的丧失。所以,在抗病育种中,人们越来越重视具有一定水平抗性品种的选育。现阶段,我国已经推广种植的抗枯萎病品种,几乎多属于此类型,故能比较长期地、并在不同生态地域均保持着较高的抗病性,如川52-128。
自Van Der Plank(1963)提出垂直抗性与水平抗性概念以后,也存在着不同看法。如Wolfe(1972)认为寄主的抗病性从感病到免疫是一个连续的统一体,病原菌的致病性,从无致病力到强致病力,也是一个连续的统一体。寄主和病原物的相互作用,可有各种各样的表现形式。Arnold等(1968)认为垂直抗性和水平抗性并没有根本的区别,只不过是总抗病性系统中的特殊情况。Zadoks等(1977)则认为,在垂直抗性和水平抗性的情况下,寄主的抗病力和病原菌的致病力间都有相互的特异作用。在与农作物病害的斗争中,有80%以上是靠抗病品种来防治或减轻其为害的。但实践表明,抗病品种在生产上应用若干年后,常会丧失抗病性。为了克服品种抗病性过早丧失,有些学者提出持久抗病性(durable resistance)或稳定抗病性(stable resistance)的概念。这是指适于某种病害发生流行的环境条件下,某一个抗病品种在大面积生产上推广多年后,抗病性仍较长期保持而不丧失,当然不意味着永久不变异。
(二)基因对基因假说
这一理论观点由Flor(1974)提出。
病原物与寄主植物的关系,即现在所提的互作关系,就是在一定条件下,植物发病过程中寄主和病原物相互作用,由一系列的生理、生化和遗传调控过程而决定病症表现的类型。Flor(1947)曾以亚麻锈病(
二、抗枯萎病育种的基本程序
棉花抗枯萎病育种的基本程序包括4个环节:①发现和创造变异;②稳定和选择变异;③鉴定和比较变异;④保持变异。所有环节都是围绕着变异进行的。
(一)发现和创造变异
棉花的性状变异可分为两种:一种是可遗传变异,即这种变异性状可以传递给以后的世代;另一种是不可遗传变异,即这种变异性状只在当代表现,不能传递给以后的世代。可遗传变异又可区分成两种类型:一类是有益性状变异,即符合育种目标的遗传性状变异,也叫符合人类栽培利用目标,能够提高经济利用价值的变异;另一类是有害性状变异,即不符合育种目标的遗传性状变异,是同人类经济利用的目标方向相反的变异。育种所需要的仅仅是可遗传的有益的性状变异。
棉花育种的任务是将现有推广品种还不具有的而生产上又迫切需要的一些新的经济或农艺性状引入新品种中,或将尽可能多的有益的经济或农艺性状集中到一个新品种中。育种的创新意义即在此。这些有益的经济或农艺性状,须是可遗传的有益的性状变异,这是育种的前提。如果没有这些变异,就不会有育种。这些有益的变异可能存在于现有品种或过时品种群体中,但更多地存在于极为丰富的种质资源中,拥有大量丰富的种质资源材料是棉花育种的基础。因此,卓有成效的育种一般需要形成一个丰富的种质资源库,育种家们深入细致地研究这些种质资源,从中寻找、发现和发掘与育种目标相符的新的性状变异。如局限在已有材料中,则很难发现符合育种目标的性状变异。
棉花育种可利用的变异主要来自两种途径:一是自然变异;二是人为变异。自然变异的引发,大多数是由于天然杂交,但也有少数来自偶然的某种外力或个别棉株自身某些不明原因诱发的自然突变。人为变异主要是通过有目的的人工杂交,但也可利用物理化学等手段,或自然界其他条件
1.自然变异
棉花是常异花授粉作物,存在一定的异交率。异交率的高低取决于传粉媒介——昆虫的种类和种群的大小。田间的传粉媒介多,棉花的天然异交率就高,容易引发较多具有变异性状的变异株,就提供了育成具有优良性状新品种的机会。这是棉花系统选择取得成功的原因。但随着棉花生产的发展,棉花害虫种类日趋繁多,为害程度日益严重,种植棉花不得不频繁地采用杀虫药剂。在防治害虫的同时,也杀死了棉田传粉媒介,大大减少了棉花异交的机会,从而降低了性状变异几率、包括优异性状变异和优异株出现的频率。这也许是多年来单纯依靠系统选择难以育成有突破性新品种的主要原因。
棉花群体中也常会出现个别的自然突变体。这主要是由于染色体畸变或某些基因位点突变而产生的变异性状。其中,有有利的,也伴有不利的经济性状变异。但发生这类突变的几率一般很低,且以质量性状为多。
2.人为变异
人为变异主要是通过人工杂交,引起基因交换、重组而发生新的性状变异。从本质上讲,这是天然异交的延伸和扩大,弥补了自然杂交几率日益低下的现状。它比之自然杂交的优点是:一能有目的地选择性状变异的方向;二能主动掌握性状变异的频率。但它的不足之处:一是不能确保有较高的成功率;二是仅限于在现有种质基因库范围内引发变异。所以,必须加强种质资源的收集、扩大和研究等基础工作,并进行较大量的杂交。
人为变异的另一途径是通过物理或化学等手段诱发新的性状变异。其优点可超越现有的棉花种质基因库,创造出新的性状变异;但缺点是:①难于诱发有利经济性状的变异,而更多的属于不利经济性状;②诱变的频率低。因此,棉花育种利用这类性状变异的难度较大。
应用生物技术,导入外源DNA,是定向诱发棉花产生新的性状变异的现代高新技术。优点是使常规人工杂交所不能跨越的亲缘障碍成为可能,在棉花染色体上不仅可导入与棉花非一个属、科、目的植物DNA,甚至可
随着中国宇航技术的发展,已多次运用太空卫星,搭载棉花种子,通过太空失重、宇宙线照射、温度和时律的变化等因素引发基因变异。但其变异方向与遗传性正在进一步研究中。
(二)稳定和选择变异
不管是自然变异或人为变异,其中,有符合育种目标需要的变异,也有不符合育种目标的变异。一般是在发现有利变异的同时,也可能相伴产生一些不利变异。这种变异,有的只是表现型的,其遗传性尚未稳定,还不能作为育种目标性状固定下来。所以,在发现或创造变异之后,育种的第二个环节就是稳定和选择变异。
稳定变异是为了保证变异的有效选择,是选择变异的前提。稳定变异的主要手段是加代。随着世代的增加,杂合体变异性状得到分离和表现型变异性状得到纯合,使所有的变异,不管是有利的还是不利的,其遗传性相对稳定下来。棉花经济性状的遗传率高低不一。一般地,遗传率高的性状,如单基因控制的质量性状,低世代时就能达到相对稳定;遗传率低的性状,如多基因控制的数量性状,只有在较高世代时才能达到相对稳定。变异性状只有达到相对稳定,不再出现明显分离时,选择才有效。不同变异性状选择的最适宜世代并不一样。为了增进加代速度,缩短育种年限,在我国,利用18°N上下的海南岛南端冬季气温较高,又是旱季的有利条件,进行秋播、冬长、春收。这样,棉花就能在一年内完成两个世代周期。但海南岛与大陆棉区的气候、生态等条件差别很大,一般在海南加代期间不进行选择,但也有加代选择成功的例证。有条件的也可在当地,冬季利用大型可控温室进行少量材料的加代。随着生物技术的发展,可利用单倍体培养技术,将变异性状的染色体加倍,或克隆复制变异株的体细胞,经组织培养,形成再生植株,获得较为稳定的变异材料。
选择变异是贯穿新品种选育始终的重要环节。棉花育种的创造性,主
(三)鉴定和比较变异
根据确定的育种目标,选择已经相对稳定的变异后,需根据留优汰劣的原则,通过科学的鉴定和比较,才能确认某些优异变异性状成为育成新品种的属性,这是选择的继续。
鉴定是在特定条件下,对某一变异性状进行有效性的直接鉴别和确认。例如,抗枯萎病性、抗黄萎病性、抗棉铃虫性、抗棉蚜性、抗旱性等。抗枯、黄萎病性鉴定,原则上是在人工接种、发病均匀的病圃中进行,也可在重发病区选择发病均匀的自然病圃中进行。抗虫性鉴定需在人工隔离环境中接种一定量的害虫数。抗旱性鉴定需设置遮雨和防止地下水浸入等设施。有些性状也可利用生物、理化反应法进行间接鉴定,但最后仍需进行直接鉴定。鉴定时要求设置抗性和感性两个对照,还需要多点、多年或多次重复,尽量减少误差,以避免年份和环境引发的影响干扰。
纤维品质测定,是确保棉花新品种纤维品质必不可少的鉴定内容。利用HVI系列测试仪器,由于每份测试样本量小,要求用随机法抽取皮棉样本,以增加样本的代表性。
育种材料的比较这一程序既重要,也繁复。随着育种进程,对照育种目标,全面考虑丰产性和有关经济、农艺性状的要求,从初级到高级,进行比较试验和鉴定,并根据结果进行逐级淘汰。对保留的材料要求要高,必须重视性状的综合表现;淘汰材料要慎重。过去的育种实践中,从原来因疏忽被淘汰的材料中,后来又选育出较突出的新品种的育种事例也有。
棉花育种材料的比较,一般从株行试验,到株系试验、品系试验、区
(四)保持变异
优良变异性状的稳定是相对的,而得到的稳定性状发生再变异则是绝对的。再变异的方向不能确定,往往是优变的几率较少,而劣变的几率更多。这就是品种的退化。
棉花良种退化是困扰棉花生产的一大难题。常常是一个良种推广不久就发生退化,削弱了良种的作用。所以,重视和切实采取有效技术,保持育种目标所要求的变异性状,就显得十分重要。这就是良种繁育。
良种繁育的任务是保持住优良品种的种性、纯度,即保证良种的品种品质。纯度是指品种纯度,并非遗传纯度。从农业生产的实际需要出发,并不要求品种在遗传上的纯化。品种本来就是一个遗传复合体,诸多性状不需要、也不可能达到遗传上100%的纯度。但必须保证主要经济性状表现型的相对一致性和稳定性。要达到这一目的,在技术上必须最大限度地避免育成的新品种发生生物学混杂和机械混杂。因为混杂的发生,必然会导致主要经济性状的退化。棉花品种的退化,往往会出现绒长变短、衣分下降、纤维变粗、铃重变轻、营养生长过旺等非人们植棉所企求的性状。
在棉花进化过程中,存在着两种选择的激烈竞争。一是自然选择,另一是人为选择。自然选择的方向是按照有利于棉花物种自身的生存和繁衍更多的后代进行的。人为选择的方向是按照人们植棉所企求的经济性状进行的。两者虽有共同点,而在经济性状方面多数是反方向的。若竞争结果是前者超过后者,即出现“退化”现象。所以,“退化”是人们从植棉目的的角度给予的评价。但从棉花物种发展的角度评价,这种“退化”恰恰正是棉种自身需要的“进化”。生物学混杂和机械混杂给人们认为的“退化”创造了条件。所以,只有一方面尽量限制生物学混杂和机械混杂的机会,另一方面当人为的选择压超过自然选择压时,才能保持种性,即保持住育种目标所企求的变异性状相对稳定,不发生劣变,不发生“退化”。这就是良种繁育的功能。
三、枯萎病抗性遗传
棉花对枯萎病抗性的遗传,国内外的研究报道较多,但结论不完全一致,多数认为,抗枯萎病性是由多基因控制的。Fahmy(1927)用免疫品种与感病品种杂交,F
Campagnaco(1971)认为,大多数杂交组合的抗枯萎病性是单基因的不完全显性遗传,F
20世纪70年代,陕西省棉花研究所、原北京农业大学(现中国农业大学)、江苏南通地区农业科学研究所等从大量杂交后代的分析中认为,杂种后代对枯萎病的抗性,与亲本的抗性水平密切相关。在感×感组合中,后代的抗性差;双亲抗性中等或在抗×感组合中,后代多为耐病;在抗×耐或抗×抗组合中,后代的抗性强等。并从育种实践中,总结出一些抗枯萎病性遗传的趋势。据王远等(1980)研究,陆地棉抗枯萎病受显性基因控制,双亲之一具有抗枯萎病性,其后代则表现抗病,因此,采用耐病、丰产、优质品种作母本,高抗枯萎病品种作父本,以培育优质、丰产、抗病品种。
校百才(1985、1988、1992)多年采用不同组合的试验表明,抗枯萎病性的一般配合力方差大于特殊配合力方差,说明抗枯萎病性的遗传中,加性效应是主要的。其狭义遗传率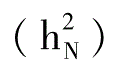 分别为42.6%、19.5%和46.5%;广义遗传率
分别为42.6%、19.5%和46.5%;广义遗传率
校百才(1985、1988、1992)多年采用不同组合的试验表明,抗枯萎病性的一般配合力方差大于特殊配合力方差,说明抗枯萎病性的遗传中,加性效应是主要的。其狭义遗传率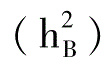 分别为42.6%、19.5%和46.5%;广义遗传率
分别为42.6%、19.5%和46.5%;广义遗传率
校百才(1985、1988、1992)多年采用不同组合的试验表明,抗枯萎病性的一般配合力方差大于特殊配合力方差,说明抗枯萎病性的遗传中,加性效应是主要的。其狭义遗传率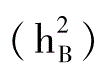 分别为42.6%、19.5%和46.5%;广义遗传率
分别为42.6%、19.5%和46.5%;广义遗传率
对棉花枯萎病抗性的遗传研究,以前多采用Griffing的配合力模型及其分析方法估算配合力效应,再用Hayman(1954)的双列分析方法估算遗传方差分量和遗传率。由于这是两个不同的遗传模型和分析方法,对一组资料分析的结果可能不尽相同,对多世代材料也不能同时进行综合分析。为此,韩祥铭等(2001)采用朱军(1997)提出的混合线性模型ADM遗传模型和ADAA遗传模型估算遗传方差和遗传率并对估算的参数进行显著性检验,从而可对亲本和组合的遗传表现进行评估。结果表明,棉花枯萎病抗性遗传,加性方差占表现型方差的49.9%,显性方差占表现型方差的13.1%,母体方差占表现型方差的0.8%,母体效应很小,主要是加性效应和显性效应,显性和环境的互作效应占表现型方差的27.0%,差异极显著,表明显性效应易受环境因素影响。广义遗传率为63.0%,狭义遗传率为49.9%,棉花枯萎病抗性遗传以加性效应为主,遗传率较高,在重病地连续选择抗病株,有利于提高枯萎病抗性。
由于不同试验所用材料、鉴定方法和技术的不同,加之棉花受多种枯萎病菌生理小种侵染,抗病基因对病菌不同生理小种的抗性不同,不同的生理小种要有不同的抗病基因,不同的生态环境,有不同的生理小种。在自然生态条件下,枯萎病菌多种生理小种可能同时存在,这样就造成了棉
棉花对枯萎病抗性与对其他病害抗性的关系,赵俊兴等(1991)对多个品种,通过线性相关分析指出,棉花品种对枯萎病和黄萎病的抗性为高度正相关,也说明品种对两种病害的抗性间有显著的相关关系,这证实选育兼抗枯、黄萎病品种的可能性。马存等(1992)的鉴定指出,抗枯萎病的陆地棉品种,也能抗苗病。Shepherd(1986)报道,棉花枯萎病的发病率与根结线虫的产卵量呈高度正相关(
关于抗枯萎病性与其他农艺性状间的相关关系,也有不同的试验结果。Patel等(1950)认为,新百万棉(NewMillion Dollar)品种的抗枯萎病性,似与不利的农艺性状相连锁,Sappenfield (1963)报道,棉花品种对枯萎病的抗性与皮棉产量呈负相关等。由此认为,抗病品种的农艺性状较差。中国在选育、推广抗枯萎病品种的初期,也有类似的观点,但后来的试验和实践证明,这种现象不是普遍规律。马存(1985)的研究指出,无论在中水肥或高水肥条件下,枯萎病越重,对皮棉产量和多数纤维品质的损失越大。在中水肥条件下,枯萎病病指每增加1,皮棉产量的损失0.94%。李俊蓝(1987)的试验也指出,枯萎病的病级与籽棉产量、皮棉产量、单株结铃数、铃重、纤维断裂长度间的
四、种质资源
种质资源或称遗传资源,是指决定各种遗传性状的基因资源。棉花种质资源包括推广品种、过时品种、引进品种、突变体材料、野生种和陆地棉种系,以及棉属近缘植物。是进行棉花品种遗传改良的物质基础,也是研究棉属遗传、起源、进化和分类的基本材料。因此,广泛、持续收集、保存、研究和利用种质资源,是棉花遗传育种研究领域中的一项基础性工作。
(一)概况
抗原是抗病育种的决定因素,而种质资源则是棉花抗枯萎病育种中抗原的来源地。我国在20世纪50年代就开始收集棉花品种和种质,并进行抗枯萎病性鉴定。1952~1953年四川省棉花枯萎病防治研究工作组收集了珂字棉8个品系及福字棉6号、鸡脚德字棉、狄胜棉、赖德福阿金等陆地棉品种(系)共33个,海岛棉品种C3173、苏-2198 两个,中棉种质有遂宁土棉、威远土棉等21个,进行抗枯萎病性鉴定。海岛棉中,苏-2198表现耐病、C3173高度感病,发病率为100%,病指79.5;中棉中,抗性最好的是仁寿紫花、盐亭小白花、三台 子花、仪陇小白花、余姚小树花、彰德土棉、遂宁28-507-47、威远土棉、宾川土棉,而玉溪中棉在田间未受棉枯萎病感染,表现高度抗枯萎病性;陆地棉发病高,但发病程度差异大,发病率为90%~100%,病情指数为59.3~71.5。
子花、仪陇小白花、余姚小树花、彰德土棉、遂宁28-507-47、威远土棉、宾川土棉,而玉溪中棉在田间未受棉枯萎病感染,表现高度抗枯萎病性;陆地棉发病高,但发病程度差异大,发病率为90%~100%,病情指数为59.3~71.5。
20世纪70年代,我国的抗病育种发展较快,对棉花品种资源的收集、抗性鉴定更加重视。1976年,陕西省农业科学院植物保护研究所采用陕西泾阳菌系,第一次对中国2238个棉花品种资源进行了苗期抗枯萎病性鉴定,结果是,无症状免疫类型22个,占总数的0.98%;高抗类型137个,占6.12%;感病类型1724个,占77.03%;其余为耐病类型355个,占15.96%。陆地棉中,表现抗性强的品种有川52-128、川57-681、陕棉4号、陕棉401、陕棉9号、川62-200、川抗病洞庭棉、晋68-389、晋68-400、云112-3、86-1号、陕棉8号、陕棉10号、咸73-74和咸73-145。中棉的抗枯萎病性强、抗源丰富,经反复试验证明,赤木黑种、赤木白种、法库白子、中棉所6号的抗枯萎病性强而稳定。
1975~1979年连续8批鉴定了中国棉花品种资源共3710个。根据病指划分为不同抗病性反应型,属免疫类型(无症状)的23个,占总数的0.62%;属高抗类型的164个,占4.42%;属抗病类型的187个,占5.04%;属耐病类型的351个,占9.46%,属感病类型的2385个,占80.46%。从不同棉种相比较,其中,以中棉抗病性较强,陆地棉次之,海岛棉、木棉抗病性最弱(表5-1)。中棉、陆地棉、木棉和海岛棉的平均病指分别为5.34、34.29、76.4和76.85。
棉种名称供试品种(系)免疫高抗抗病耐病感病病指0病指0.1~10病指10.1~25病指25.1~50病指50.1~100个数%个数%个数%个数%个数%陆地棉3102130.42993.191374.422608.38259383.58海岛棉27841.4427498.57中棉308103.256521.104715.268627.9210032.47木棉1715.881694.11草棉5360.00240.00合计3710230.621644.421875.043519.46298580.41
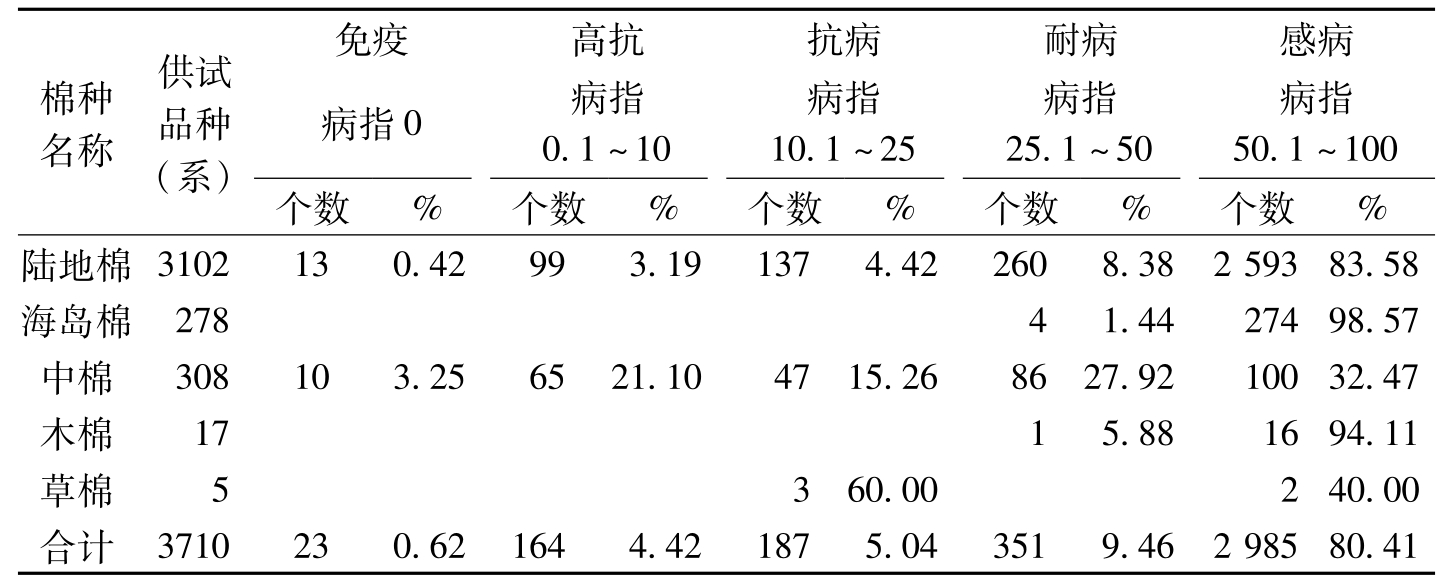
表5-1 不同棉种对枯萎病苗期抗性鉴定
1993年,陕西省农业科学院植物保护研究所又对348个亚洲棉(即中棉)品种进行抗枯萎病苗期抗性鉴定,病指为1~25的有288个,占82.8%;病指为25.1~50.0的有34个,占9.8%;病指为50.1~100.0的
(二)抗原种质52-128和57-681的育成与应用
52-128和57-681是中国育成的第一批高抗枯萎病抗原种质。四川省棉枯萎病工作组自1952年在发病90%以上的大榆区紫云乡105亩(667m
1981年选用有代表性的生理型Ⅰ号川F
这两个抗源品种育成后,于60年代初被陕西、山西等省引进试种,表现出抗性强、稳定性好,从而用作抗原进行杂交转育或取材,育出陕4号、
据《全国农作物审定品种名录》(2005)、《中国棉花品种及其系谱》(1996)、《中国棉花品种系谱图》(2000)资料分析,以及中国农业科学院棉花研究所等50余家育种单位利用这两个抗原的应用证明统计,全国利用抗原52-128、57-681育成棉花抗病品种128个。在不同转育代次育成的抗病品种中,集中来源于第二代至第五代的有121个,占94.5%;被利用作母本、父本育成的品种分别各占有54.7%和31.3%(表5-2、表5-3)。利用52-128抗原品种作抗原,通过杂交或系统选育的抗病品种,在各地方推广应用都表现抗性良好(表5-4)。
省份育种单位(个)80年代90年代2000~2004年总计河北765112山西33429辽宁244江苏9112720浙江111安徽1516江西111山东31427河南9812222湖北49211湖南21124四川5511319陕西29211新疆111合计503569128
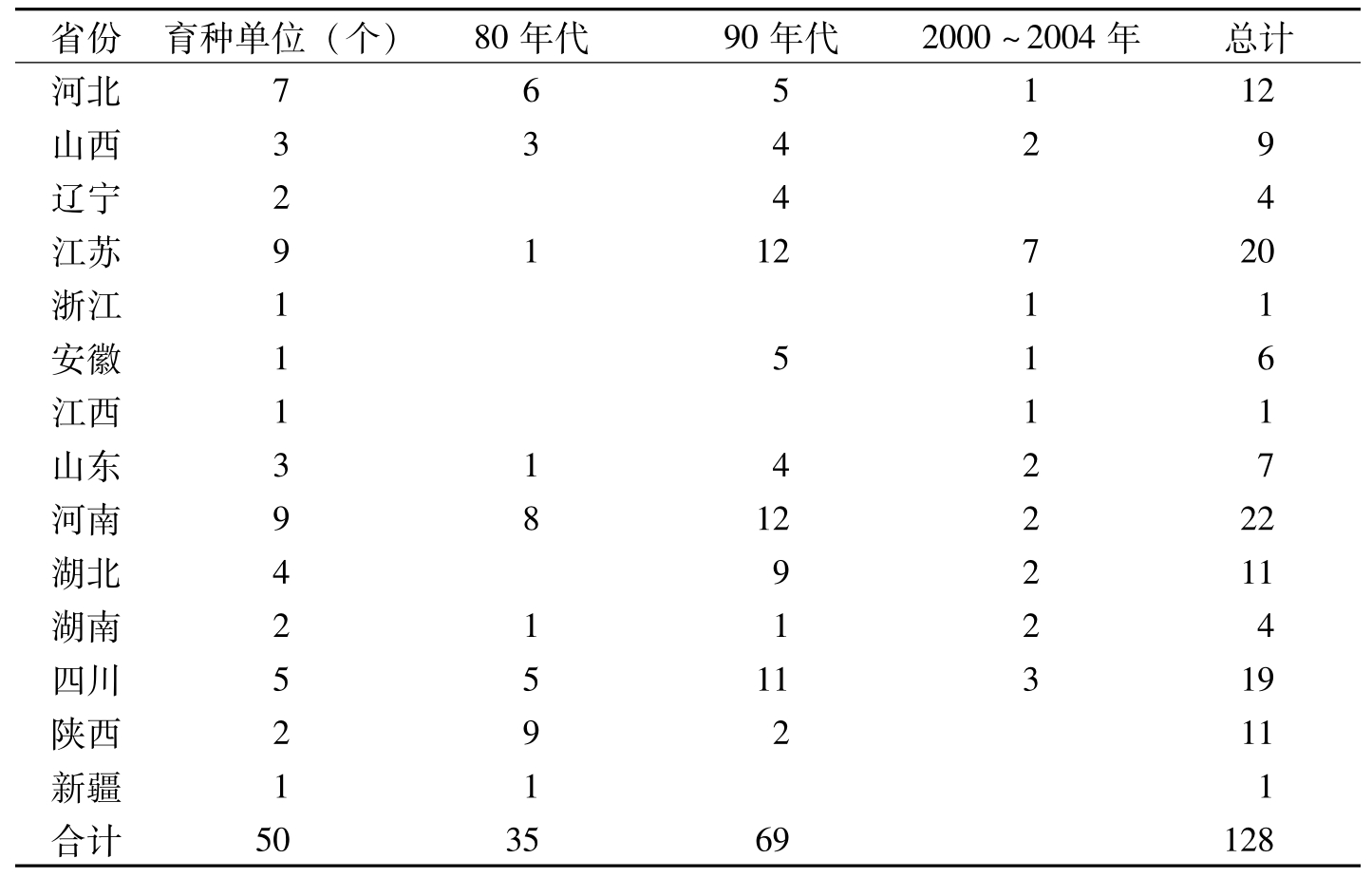
表5-2 不同时期不同地区利用抗原52-128、57-681育成的审定品种数
抗原用途一代二代三代四代五代六代总计母本(♀)11029191170父本(♂)46917440系统选育15542118合计6214340171128
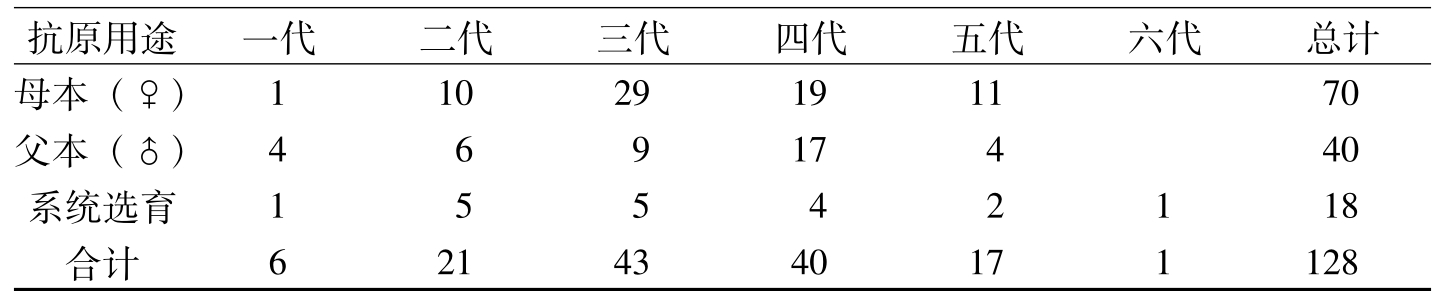
表5-3 抗原种质52-128、57-681不同转育代次育成的品种数
项目四川湖北新疆江苏云南山西陕西辽宁上海山东河南河北北京浙江甘肃安徽湖南江西合计供试菌系179985111466777552911129抗性反应高抗01600000020100020012抗病861327114546422151072耐病71043432110232110136感病2121000000100101009
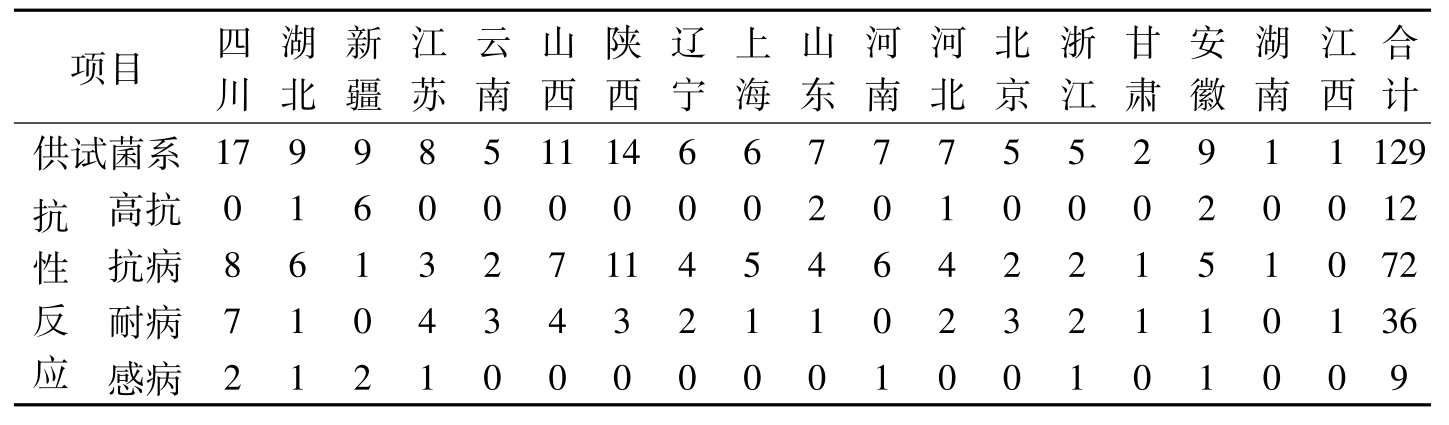
表5-4 52-128对不同地区菌系的抗性反应
抗原种质在育种上应用成败的关键,一是抗原抗性稳定性。抗性稳定而持久的抗原,转育利用的抗性不易衰失,能持久保持利用;二是抗原种质的遗传效应,这是转育利用的基础;三是抗原种质经济性状的遗传效应,这对利用价值影响极大。针对这三个关键问题,叶鹏盛等(2007)对52-128和57-681的抗性遗传做了研究。结果证明,这两个抗原的抗性长期稳定(表5-5,表5-6),且抗性遗传效应好、遗传率高(表5-7),其他不良性状在杂种后代中的遗传不显著(表5-8),为我国利用两个抗原的抗性,选育抗病品种奠定了良好的基础。
供试品种种植1年种植2年种植3年种植4年病圃无病地病圃无病地病圃无病地病圃无病地521284.504.504.503.502.752.750.752.75576816.003.507.254.502.252.251.253.25川73274.008.009.008.003.503.502.507.25川4149.005.755.758.001.001.001.250.75达棉1号30.7556.7523.556.7535.0035.0017.2536.50显著性(t值)0.53240.62710.06431.1214

表5-5 在病圃和无病地筛选后抗原的病情指数
世代52128×达棉1号57681×达棉1号达棉1号×52128达棉1号×57681病圃无病地病圃无病地病圃无病地病圃无病地显著性(t)F135.0019.2541.0024.5018.5023.7521.7524.502.5687F231.2519.2519.2524.5012.7517.2518.7516.252.3072F310.2519.7515.5025.507.7520.258.0025.508.6017**F42.0013.751.2524.508.7521.759.7520.509.0275**
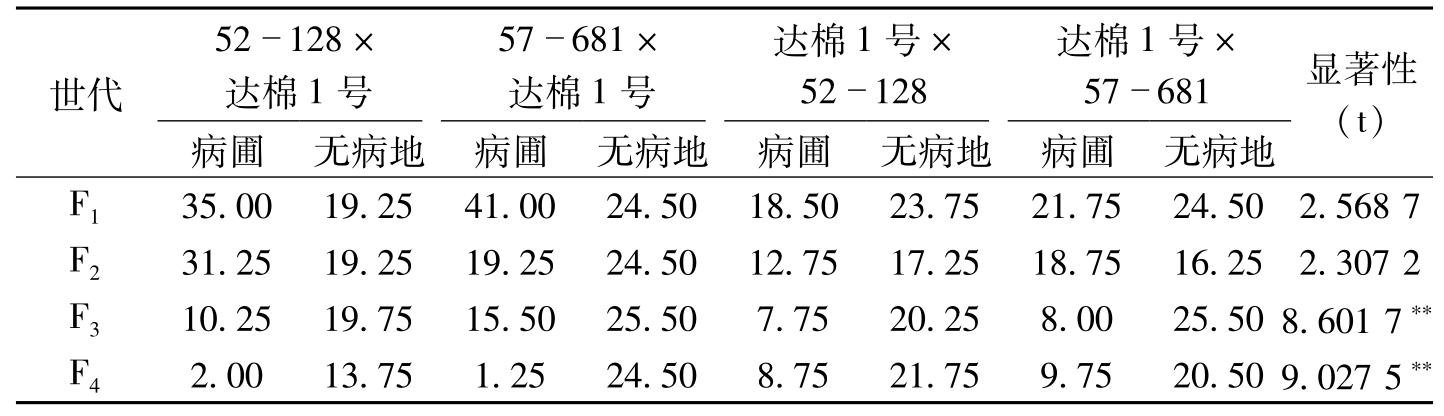
表5-6 在病圃和无病地筛选后抗原不同杂交后代的病指
抗源及组合平均病指F1F2中亲值相对优势(%)广义遗传率(%)狭义遗传率(%)52128×达棉1号29.5039.2539.75-25.7968.952.157681×达棉1号25.5034.0040.50-37.0473.156.77329×达棉1号29.0037.2541.25-29.7070.548.2川414×达棉1号29.7539.7542.00-29.1767.443.6达棉1号×5212835.2535.0039.75-11.3272.953.7达棉1号×5768134.5034.5040.50-14.8175.452.4达棉1号×732737.0038.2541.25-10.3072.353.4达棉1号×川41437.2541.7542.00-11.3169.246.5521284.00576815.50川73277.00川4148.50达棉1号75.50
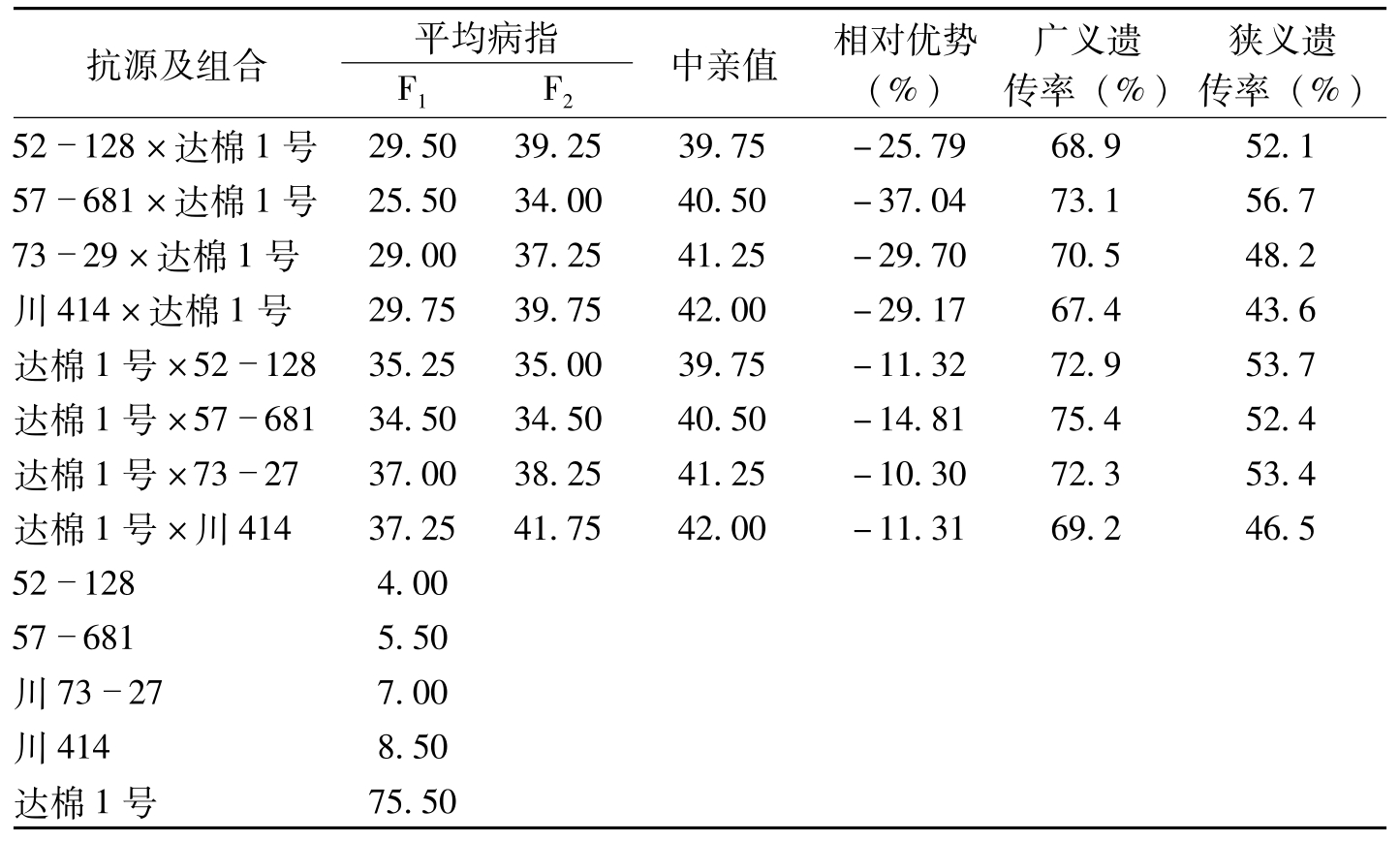
表5-7 抗原种质52-128、57-681的抗性遗传效应
抗源及组合株高(cm)单株铃(个)衣分(%)F1F2中亲值F1F2F1相对优势(%)F1F2中亲值F1F2F1相对优势(%)F1F2中亲值F1F2F1相对优势(%)52128×达棉1号99.3100.188.491.012.319.717.317.017.915.836.137.236.737.8-1.6

表5-8 抗原种质52-128、57-681的主要经济性状的遗传表现
抗源及组合株高(cm)单株铃(个)衣分(%)F1F2中亲值F1F2F1相对优势(%)F1F2中亲值F1F2F1相对优势(%)F1F2中亲值F1F2F1相对优势(%)57681×达棉1号93.897.685.186.010.221.018.916.118.930.438.438.337.538.8-2.4川7329×达棉1号86.790.083.483.83.823.220.320.119.715.440.340.239.840.31.3川414×达棉1号85.189.782.784.82.919.418.919.818.8-2.039.840.140.340.3-1.2达棉1号×5212898.996.388.491.011.923.218.817.217.952.837.839.636.737.8-3.0达棉1号×5768187.188.885.186.09.424.621.216.118.934.938.938.337.538.83.7达棉1号×732785.287.683.483.82.323.722.620.119.717.940.941.239.840.32.8达棉1号×川41490.385.182.784.89.221.719.819.818.89.640.140.040.340.3-0.4
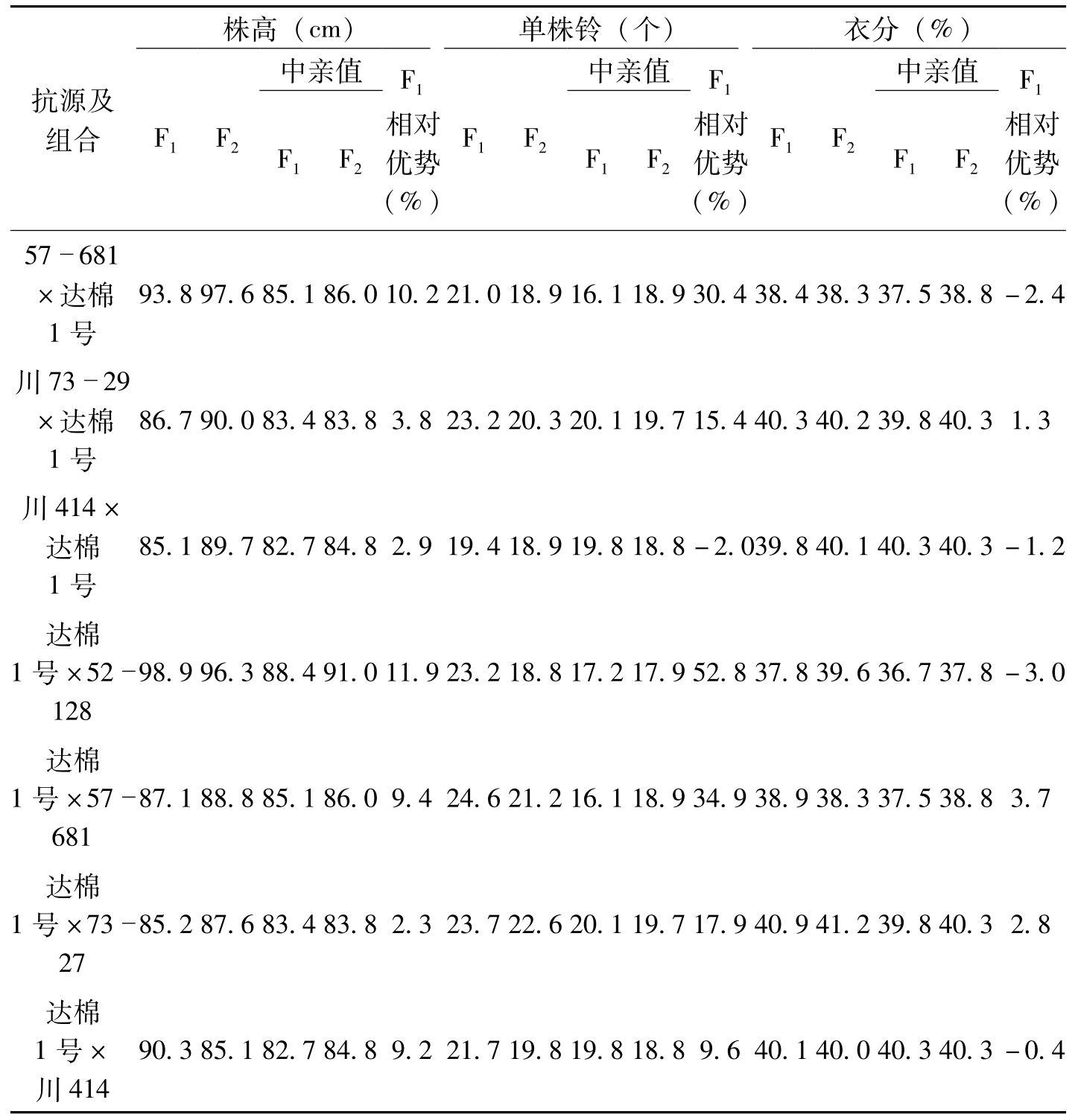
表5-8 抗原种质52-128、57-681的主要经济性状的遗传表现(续)-1
(三)种质资源创新
磷作为植物生长发育的必需营养元素之一,不仅是植物体内许多重要化合物的组分,而且还以多种途径参与植物体内各种代谢过程,在人类赖以生存的生态系统中,起着不可替代的作用。植物吸收磷的主要形式是 ,它们在土壤溶液中的浓度很低,一般只有1.5μmol/L。因此,磷容易被土壤固定,在土壤溶液中的移动性很差。大多农业土壤因长期施用磷肥已成为潜在的磷库,磷肥效应与土壤有效磷含量呈负相关,
,它们在土壤溶液中的浓度很低,一般只有1.5μmol/L。因此,磷容易被土壤固定,在土壤溶液中的移动性很差。大多农业土壤因长期施用磷肥已成为潜在的磷库,磷肥效应与土壤有效磷含量呈负相关,
陆文龙等(1999)研究表明,营养高效基因型是指在缺磷条件下,能够利用自身根系分泌的有机酸活化土壤中难溶性磷,协调生长代谢,维持正常的生长发育过程,最终形成较高产量的基因型。由于植物对磷素的利用能力具有模糊性,耐低磷基因型和非耐低磷基因型的表现特征是连续的,没有明显的界限,磷素的影响不仅表现在相对干物重上,而且与植株的地上部鲜重、叶绿素含量、总叶面积等性状也相关。王士杰等(2009)首次在确立的筛选指标基础上,从88个棉花抗枯、黄萎病品种中筛选出25个耐低磷基因型品种(包括中棉所9号、中棉所15、中棉所21、中6331、86-1、豫棉1号、豫棉8号、豫棉22、冀棉7号、冀棉20、冀合3028、鲁无401、鲁棉14号、陕棉11、苏棉12、徐261、泗棉3号、川棉109、绵阳83-21、辽棉12、皖棉11、GK1、Stoneville 603、Deltapine 61和渤棉抗4)和3个耐低磷极端基因型品种(中棉所21、中99和陕棉11)。这些品种可以作为培育耐低磷胁迫的棉花抗病品种以及开展棉花耐低磷QTL定位与克隆等研究的重要资源。
鉴于新疆长绒棉枯萎病迅速蔓延,给长绒棉生产带来巨大威胁,生产上迫切需要长绒棉抗病品种来减轻枯萎病的为害。在长绒棉现有的品种和品系中很难找到抗源。为此,武刚等(2006)利用抗枯萎病的陆地棉品种作种间杂交选育,经病圃多年筛选、鉴定出4个高抗枯萎病,而纤维品质达到长绒棉标准的新种质。
(1)237-1
生育期143天,绒长36.5mm,比强度31.8cN/tex,马克隆值3.7,单铃重2.4g,衣分31.8%,皮棉产量85.7 kg/667m
(2)237-4
生育期146天,绒长36.5 mm,比强度37.0cN/tex,马克隆值3.3,单铃重2.5g,衣分32.8%,皮棉产量90.6 kg/667m
(3)240-2
生育期148天,绒长36.6 mm,比强度30.4cN/tex,马克隆值3.7,单铃重2.5g,衣分32.3%,皮棉产量82.3 kg/667m
(4)240-4
生育期147天,绒长37.3 mm,比强度33.0cN/tex,马克隆值3.7,单铃重2.3g,衣分29.4%,皮棉产量78.6 kg/667m
刘泽辉等(2008)对上述4份新种质在进一步杂交转育研究后认为,所选育的新品系同抗病新种质的抗病性没有明显差异,年度间抗病性差异小,说明抗病性呈显性性状、能稳定遗传。这为今后长绒棉抗枯萎病育种提供了很好的抗源材料,为棉花产业发展奠定了基础。但同时也指出,长绒棉抗枯萎病新种质在田间长势旺、抗病性强,但铃重偏低,产量不太理想。此外,这些高抗枯萎病新品系种植在多年陆地棉黄萎病很重的棉田,偶尔发现一两株有黄萎病病症的棉株,也许是杂交后代具有一定的陆地棉基因。因此,新种质抗性基因遗传特性、抗性生理生化方面的研究以及产量的提高等还有待进一步加强。
五、常规育种方法
在制定抗病品种育种目标时,既要求选育的品种具有抗病性,同时又要丰产和纤维品质优良,即除抗病性外,其他性状应与当地推广的非抗病丰产品种要求相似。事实上,一个耐病而丰产优质的品种往往比高抗而低产劣质的品种更有实用价值。进行抗病育种时,要注意不仅培育只对某一种病害的某一生理小种具有特异性抗性的品种,而且要培育多抗(水平抗性)品种,因为具有特异性抗性品种在出现新的菌株和生理小种时,抗性即消失。棉田单一的枯、黄萎病区较少,多为混生病区,因此,培育具有兼抗特性的品种,在制定抗病育种目标时也应加以考虑。
(一)系统育种
在自然界棉花群体中,个体间总是存在着微小或明显的性状差异,包
1.一般原理与方法
系统选育方法能在抗病育种中取得一定效果的原因在于:①棉花是常异花授粉作物,由于经常的天然杂交,增加了遗传基础的复杂性。一个棉花品种,一般人们较重视的一些农艺性状和经济性状是相对一致的,而人们没有注意的或缺少一定条件暂时未表现的性状(如在无病地的棉花抗性),个体间存在差异,有时差异可能很大。如能为棉花创造一定的条件(如发病条件),就可使个体间抗病性差异表现出来。重病地提供了筛选的条件,因此,一次选择可能选出抗病的个体。②棉花对枯萎病的抗性遗传是由基因所控制,当一个品种处于一种复杂群体状态时,这些抗病基因可能分别存在于不同个体上。在经过一次抗性选择的基础上,由于天然杂交,杂合体抗性基因分离重组,必然会产生抗性基因积累,连续选择便有可能把具有多个抗性基因累加效应的个体选择出来,从而进一步提高了抗性。③根据Vidhyasekaran(1988)“几乎所有植物都存有抵御微生物入侵的防卫机制”的论断,寓意着植物的抗性是固有存在着的。从抗性的性质来看,植物的抗病机制包括预存性被动机制,以及被病菌侵染后激发产生的主动防卫机制。在主动抗病性中,有两套基因先后起作用,即抗病基因和防卫反应基因。抗病基因产物对病菌无毒基因产物有识别作用,从而可诱导防卫基因表达,产生一系列防卫机制,所以,抗病基因产物有识别作用。抗病基因是一种效应分子(抗病蛋白),本身无杀菌作用,真正起作用的是通过防卫机制产生的物质,在形态和生理生化上的许多改变,这些抗病变化包括,植物保护素的合成、细胞壁修饰(愈伤组织、木质素和酚类化合物在壁上沉积)、富含羟脯氨酸糖蛋白的积累、蛋白酶抑制剂和能抵御病原物细胞壁的水解酶(角质酶和葡萄糖酶)的产生。所以,在植物抗病性反应中包括两个阶段。第一阶段为决定阶段,通过病菌无毒基因产物和寄主抗病基因产物相互识别决定寄主抗病性表达;第二阶段是表达阶
在中国棉花抗病育种史上,系统育种是20世纪80年代前所采用的主要育种方法。据1974年统计,全国育成的148个品种中,67.6%的品种是采用系统育种法育成的。70年代所育成的48个品种中,也有26个是采用系统育种法育成的。川52-128和川57-681是我国棉枯萎病的主要抗原,两者都是在重病田上经连续选择而育成的。川52-128选自古老的栽培品种德字531,川57-681选自岱字棉15,86-1来自陕65-141,陕3563、川73-27、鲁抗一号均选自陕4。中棉所3号选自乌干达3号。
系谱选择法是系统育种中最常用的方法,它不仅用于从自然变异选择的系统育种,也应用于杂交、化学和物理方法诱变等人工方法产生的变异群体。在自然变异和人工创造的变异群体中选择单株或单铃种成株行或铃行,每株行或铃行为一个家系或“系统”,由于“系统”间差异比个体间差异大,故先选择优良系统,而后在当选系统内继续选株或选铃,种成下一代株行或铃行。选择连续进行数代,等系统内植株性状已趋一致时,选留符合育种目标的系统,按系统收获。翌年升入设有重复的鉴定圃比较产量,通过一年或多年、一地点或多地点产量比较,鉴定出产量高并具有育种目标要求的品质和抗性的系统繁殖成为品种。
系统育种方法简便,收效快,是棉花育种的主要方法之一。但该法也有一定的局限性,表现在:
①系统育种的效果主要决定于某品种群体的遗传变异性;而其遗传变异性,表现在天然杂交,基因突变及剩余变异等天然变异。一般地说,这种天然变异几率不会太高,而符合育种目标所要求的有利变异率更低,因而选择效率不高。
②应用连续单株选择育成为品种是由一个单株繁衍而成的群体,其遗传基础较窄;对环境条件的适应力差,改进提高的潜力有限。前苏联育种家1978年的试验指出,由21个优良株系组成的品种C-4539群体,经一
为了提高系统育种的效果,可采用下述方法加以改进。
①有重复的株行(系)试验。现代的育种方法应能保持和控制遗传变异,即能使育种家容易判明育种材料的遗传变异和由环境条件引起的非遗传变异;而且一个群体对选择的潜在反应的利用决定于所应用的技术方法的有效性。假如所应用的方法不是很敏感的,那么,大多数的遗传变异不仅不能被发现,而且会由于同质性的增加而丧失。
由于一个单株后代的种子数量少,故对其后代进行试验鉴定时,增加重复次数比扩大小区面积更能充分利用有限的种子和提高选择的可靠性。
②混系法。在改良品种时,为了保持其遗传可塑性,可按一定标准将若干个在形态上基本相似而又不完全相同的家系,合并成为混系品种,其效果比单一家系品种较好。
用系统育种方法选育抗病品种,须注意以下一些问题:①整个试验从选择单株到品比试验,自始至终都应在发病均匀的重病地上或人工病圃上进行。以便在表现抗性程度的基础上比较株系间的丰产性和纤维品质。②以当地推广的品种作对照。③除进行一般生育期调查外,应在苗期、现蕾后、结铃盛期以及收获前(劈秆)等不同时期进行发病情况调查。④株行试验可侧重观察抗性,参考结铃性和其他农艺性状的表现。品系预备试验和品种比较试验,则应在表现抗性的基础上注意丰产性和纤维品质。表现较突出的材料可提前进行多点试验,同时进行纤维品质的鉴定,以加速
2.抗性定向培育法
这一方法由高永成等(1995,1996)提出。应用这一方法,徐州地区农业科学研究所、中国农业科学院棉花研究所等单位,从国外新引进的原来感病品种,于5年时间内,改造成抗病品种获得成功。如从1979年开始试验,徐州地区农业科学研究所培育的徐州142、中国农业科学院棉花研究所培育的中棉所7号及西北农业大学培育的西农3195等原来都是高产感病品种,经过5年左右的病床病圃连续群体选择法,徐州142病床枯萎病病株率由原来的97.4%降低为8.5%,岱字棉16病床枯萎病病株率由原来的91.6%降低为8.5%,中棉所7号由原来的99.9%降低为11.3%,西农3195由原来的95.9%降低为7.5%。当选材料,不但由原来感病的转变为抗病的,并且由于连续群体选择,不仅仅着眼于抗性,同时兼顾产量、产量因素及品质性状的同步提高,因而,能将原来的感病品种有效地转变为抗病而综合优良性状的新品种。高永成等(1995)和张云青等(1997)对这一方法作了理论上的分析,认为:
①棉花作为枯萎病病原菌的寄主确有在变异的生存条件下,即病茵连续侵入并增大压力条件下,逐渐而又明显地改变自身遗传性以适应新的环境能力。生物的适应变异现象确实存在。同时,棉花寄主确有通过后天获得性遗传累加作用,逐年逐代增强抗性。最终由原感病品种经强化成为高抗品种。没有适应性的变异和后天获得性遗传的累加相互作用,抗性增进不可能,这是抗性提高和定向培育的根本认识之所在。
②原感病品种对毒素处理反应迟钝,出现萎蔫现象时间晚,但萎蔫指数高。抗性转化年代愈久、抗性愈高的品种对毒素处理反应愈敏感,出现萎蔫时间也愈早,但萎蔫指数愈低,甚至出现恢复现象。电镜解剖观察到原感病品种导管褐变时间晚,对毒素处理反应迟钝。导管褐变后出现保护性物质——侵填体少,比率小。增进抗性后,导管的褐变现象出现时间,
③原感病品种过氧化物酶活性与多酚氧化酶活性高,而抗坏血酸氧化酶活性极低(0值)。定向培育年限愈长,抗性愈高的品种过氧化物酶活性和多酚氧化酶活性愈低,而抗坏血酸氧化酶活性愈高,表明感病品种和抗病品种的生化代谢型确有差别。前两种酶及活性与感病性有关,后一种酶及活性与抗病性有关。抗性转化的实质即代谢型的转化。
④育成的抗病品种与感病品种棉籽粉用SDS电泳法进行蛋白质分析表明,感病型品种β组分色谱带量少色淡,抗病品种β组分带量多色深。β组分蛋白质估计为球蛋白。说明抗性转化过程中棉株生化代谢型转化,会影响到种子生化代谢型的变异,这是棉花抗性品种与感病品种生化物质成分的差异。种子球蛋白用SDS-PAGE法分析发现感病的岱字棉16种子球蛋白由10种分子量不等的蛋白质组成,而岱字棉16则由8种蛋白质组成,前者含有较多高分子量蛋白,后者含较多低分子量蛋白,表明抗性增进过程中,种子形成高分子量球蛋白质合成减少、低分子量球蛋白合成增多。
抗性定向培育法的关键技术为:①培育的对象应是需要注入和强化抗性的农艺性状优良品种;②注重培育病床、病圃,强化菌种分离与培养;③病床、病圃发病高峰期,强化对棉苗(株)的选择、严格淘汰;④纤维品质测定、淘汰与选择;⑤经3~5年严格筛选,同时,择优进入继续强化抗性的良繁体制。
为探讨定向培育法通过病圃加压筛选获得的抗病品种(系)提高抗枯萎病性的病理学机制,吕学莲等(2008)以病圃定向筛选抗枯萎病大幅度提高的棉花材料和原感病品种为对象,研究接种枯萎病菌后抗、感材料的病理学变化。结果表明,两者在宏观病理学方面存在差异,感病品种接种7天开始显症,25天严重发病,整株维管束变色;抗病材料生长健康或只有轻微症状,维管束变色部位仅局限在子叶节以下。在细胞和组织病理学方面,两者也存在差异,侵入前期,抗病材料被侵入细胞中,有细胞壁加厚现象;侵入中期,病菌可通过表皮层侵入感病品种的薄壁组织,但只能
(二)杂交育种
杂交育种是指用基因型不同的品种(系)作亲本,通过有性杂交获得杂种,继而在杂种后代中进行选择,培育成符合生产发展需要的新品种的方法。在杂交育种中,通过人工有性杂交,可将不同亲本的优良基因组合在F
作物产量、品质、抗性等性状之间,常有负相关现象,已有许多研究者讨论过这一问题,并提出打破或缓解这种负相关的方法。Weikaivan等(1995)提出关于作物性状负相关的见解认为,这是育种上一个极为复杂的问题,比以往人们所提到的应给予更多的重视。据Donald等(1996)认为,许多与生长发育有关的性状都是相互联系的,稳定地保持正或负的相关关系。由于相互联系的性状之间的负相关,阻碍了育种的进展。认为不利性状的联系有3个主要来源,遗传连锁、一因多效,以及一定环境影响下的性状间的相互联系和补偿作用。由于遗传连锁的负相关势必延缓育种进程,但是,只要有充分大的杂交分离群体,并确切地鉴定重组型,遗传连锁是可以打破的。遗传连锁一旦打破,重组型将成为既持久又有利的遗传连锁了。与此相反,由于一因多效的负相关,既不受遗传也不受环境因子改变的影响,比较难以打破。由于环境导致互补而产生的性状间的负相关也难以打破。主要是与产量有关的性状,相互联系形成一个体系。在这个系统内,一个或多个性状必须在表现程度上有所递减,另一性状的表现程度才能有所提高。相互联系的性状应该看成一个有常数容量的生物学体
通过何种育种方法才能有效地打破这种遗传上负的关联性,获得理想的重组体,育成符合育种目标要求的综合性状优良的新品种,这是国内外棉花遗传育种家一直在努力研究的课题。
棉花育种大多采用杂交育种法,显然已取得了显著的成效,但这种方法存在显而易见的缺陷。首先是杂交次数少,不利于有利基因通过交换达到重组,不利于基因加性效应的积累,极大地限制了理想个体出现的几率;其次是难以打破皮棉产量与纤维品质、产量与抗病性性状遗传上的负相关性,难以选择出综合性状优良的个体。针对这些突出问题,国内外棉花育种家试验过多种育种方法。主要有:
1.保持基因流动性育种杂种库体系
这个体系是张凤鑫(1987)等研制成功的。
育种上不能把各种有益性状集合在一个基因型中,这是由于存在性状间的不利关系,这种关系来自连锁,或“多因一效”,或“一因多效”,但主要是连锁的作用。要从根本上解决这一问题,只有通过遗传信息序列的广泛重组才能改变其连锁状态。
把育种群体作为一个动态基因库来处理,通过多个亲本多次的相互交配(含分裂交配)及随时根据需要输入新种质,通过遗传的高度杂合化,打破原有亲本的遗传系统,促成遗传信息序列的广泛重组;打破遗传连锁,释放出潜在有益变异,再按育种目标施以相应的(病、虫、逆境、产
张凤鑫等应用该体系进行育种验证,结果表明,该体系是行之有效的,一是抗性进展快,枯萎病病指较亲本均值降低90%左右,黄萎病病指降低59%,苗病死苗率降低80%,一部分材料还兼抗棉蚜、棉红铃虫、棉铃虫,如杂种优势利用新材料高柱头系102-1,高抗枯萎病,抗黄萎病、抗棉蚜、棉铃虫,高抗红铃虫。二是铃重、单铃子数、衣指、籽指、衣分、单铃皮棉重、2.5%跨距长度、比强度、马克隆值的遗传变异分别扩大2~3倍。抗性与皮棉产量的遗传关系得到改善,产量与苗病死苗率、枯萎病、黄萎病的病指遗传相关系数分别为
2.混选混交体系
该体系由马家璋等(1987)建立。其指导思想是,通过混交,打破育种性状间的不利遗传负相关;通过混选获得优良基因的重组体。其主要步骤是,以育成的遗传基础丰富的选系为共同亲本,同时与几个各具特色的目标性状亲本杂交。F
该体系的遗传改良效果也是比较好的,表现在:①显著提高产量、早熟性、2.5%跨距长度的平均数。枯、黄萎病的病指保持在高抗和抗的水平;②扩大遗传变异幅度;③改善性状间关系,加强了产量与铃数、铃重间正相关,降低株铃数与铃重、籽指与衣分率之间负相关,皮棉产量与纤维比强度的遗传相关系数由-0.397变为+0.494,株铃数与比强度相关系数由-0.901变为+0.71,衣分与马克隆值相关系数由+0.34变为-0.411,早熟性与皮棉产量相关系数由-0.282变为+0.797。
马家璋等采用该体系育成兼抗品种中棉所23。山西省农业科学院棉花研究所采用这一方法也育成了晋棉11。
3.修饰回交体系
该体系由潘家驹等(1990)建立。修饰回交(modificd backcross)就是回交后代选系间的彼此再杂交,可以培育棉花多系品种。在棉花抗病育种中,通过回交产生若干同质系,然后将各同质系按一定要求混合组成多系品种。通过修饰回交研究,将杂种品系间杂交与回交结合,综合两者特点,回交纯合快,缩短育种年限,后代只聚合回交亲本基因型,选择容易成功。其程序是:
选用一个丰产品种作为轮回亲本(A),以优质品种(B)、抗病品种(C)为授予亲本,以品种A分别与品种B、C杂交,其后代分别以A作为轮回亲本回交,分别从回交后代中选拔优系,然后进行不同回交选系的杂交。其程序模式如图5-1所示。
通过1980~1990年两轮试验结果表明,修饰回交法对削弱丰产与抗病、抗病与纤维比强度之间的负相关程度有一定效果。第一轮试验中,皮棉产量和病指的相关系数由原来的-0.51转变为-0.21。在第二轮试验中,无论抗病株率与皮棉产量或病指与皮棉产量的直线回归分析,4个修
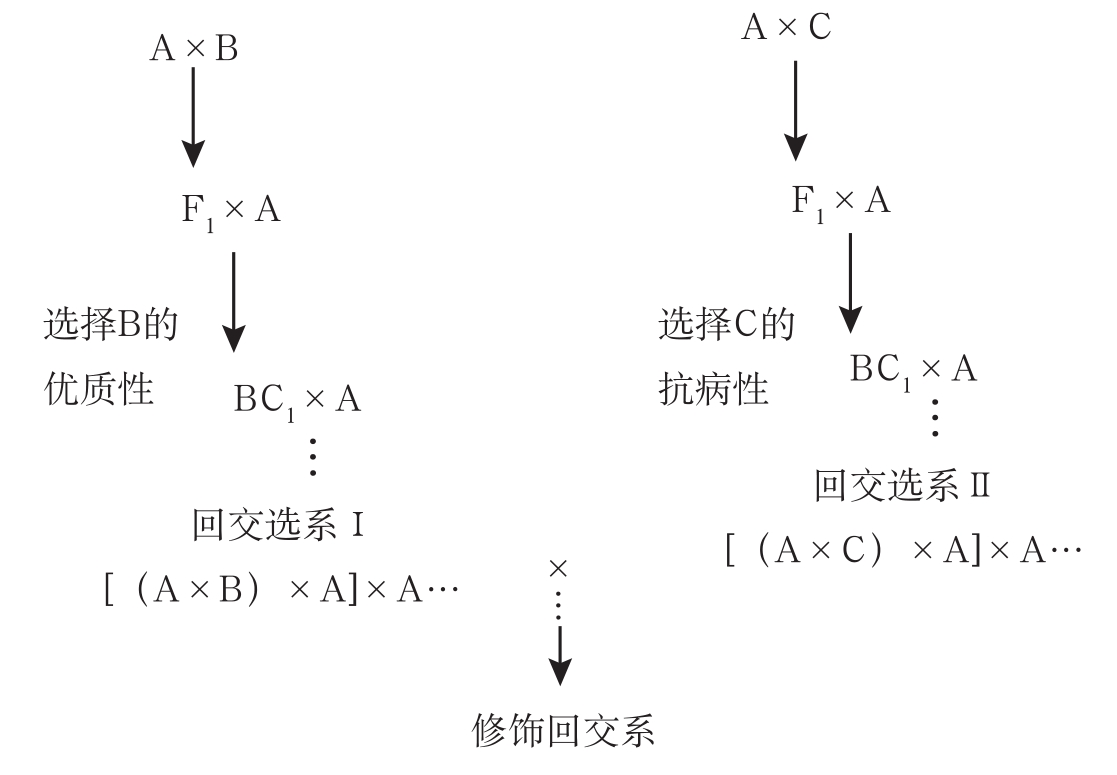
图5-1 修饰回交体系
4.MAR育种体系
MAR(Multi Adversity Resistance,多抗逆性)体系是Bird等(1980)通过研究种子状况、抗细菌性角斑病和抗性基因之间的遗传关系(图5-2)后提出来的。
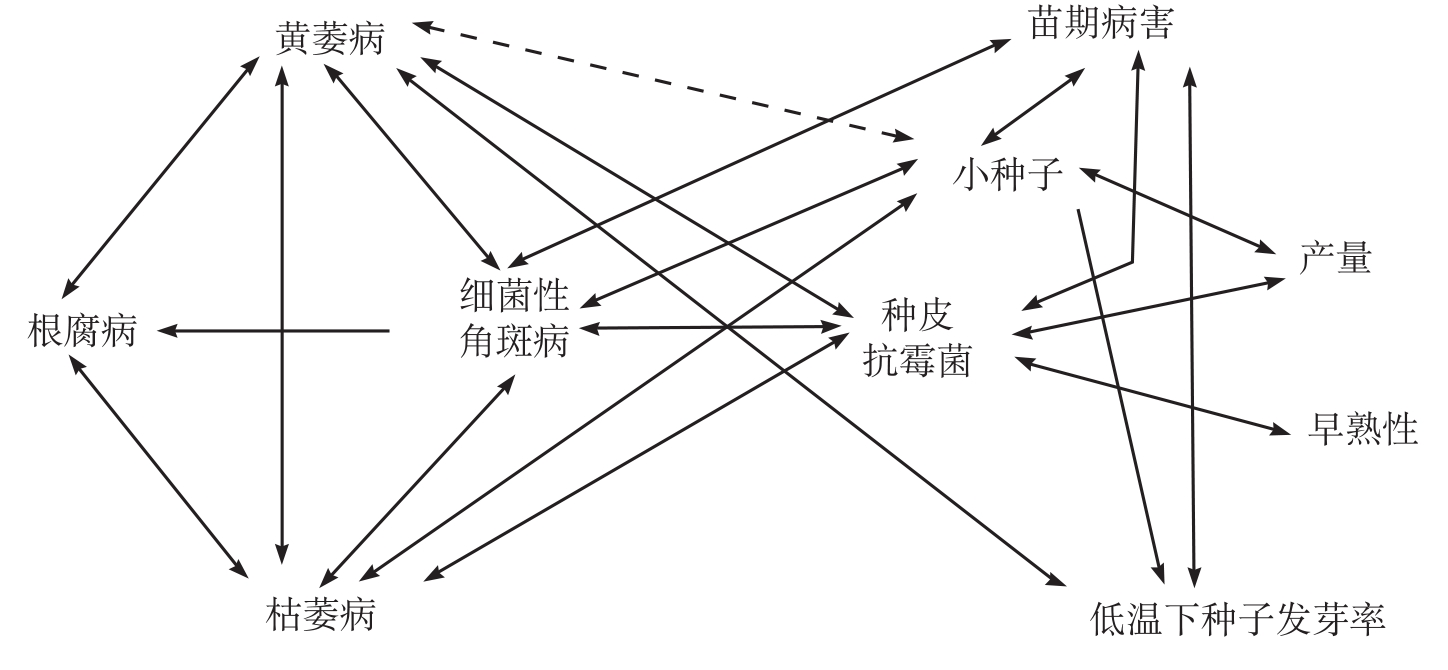
图5-2 MAR体系中抗病基因与其他性状基因之间的遗传关系
从图5-2中可得知:
①抗细菌性角斑病与枯萎病(根结线虫病复合体)之间有很强的相关关系;
②抗细菌性角斑病与黄萎病、根腐病以及种皮抗霉菌之间的相关关系较小,但显著;
③抗枯萎病(根结线虫病复合体)与黄萎病、根腐病之间也存在一定的相关性;
④在低温(13~18℃)条件下,降低发芽速度与抗黄萎病、苗期病害有关;
⑤种皮抗霉菌的特性与产量、早熟性之间有高度相关;
⑥小种子与在低温下发芽快、易感黄萎病密切有关。
根据以上研究结果,MAR体系育种方法的关键是:种皮抗霉菌,种子在13.3℃下处理8天后的发芽状况,以及子叶期对多种不同角斑病生理小种的抗性性状的选择。通过对这3种性状的选择,达到改良抗病性、提高产量和提早成熟的目的。具体的技术要点如下。
①根据育种目标,选用具有抗性、高产和纤维品质好以及成熟早的材料用作杂交亲本,配制复式杂交组合。例如,品种SP21S是选自杂交组合(SP21F×SP33F)F
②单株选择开始于复式杂交组合的F
③脱绒后的种子放入盛有1.5%洋菜培养基的培养皿内,每皿25粒。培养皿不加盖,让种子与培养基暴露在空气中,以自然的方式感染真菌孢子,主要是镰刀霉和交链孢霉。最后用带有小孔的塑料纸把培养皿封上,放进13.3℃的人工生长箱内历时8天。8天后检查种皮表面抗霉菌和发芽情况。当选种子的标准是:种子表面不带霉菌,且刚露白(胚根仅伸出种皮1mm左右)。由于不同材料抗种皮霉菌和种子发芽的能力有差异,因而在实际掌握这两个标准时,应尽可能选择种皮表面不带霉菌或少带霉菌,胚根伸出种皮尽可能短的种子。
④当选种子在温室中播于用立枯病和猝倒病原菌接种过的土壤中,5天后检查发病情况。正常苗保留,感病苗淘汰。
⑤当选的正常苗再用4种不同生理小种的细菌角斑病菌混合液在子叶背面中部接种,10天后检查发病情况。免疫的苗(接种伤口处不发黑或没有向附近健康组织蔓延)保留,其余的淘汰。同时,检查每个幼苗的下胚轴部分(茎与土面交界处),感病发黑的淘汰。
⑥当选的幼苗即为MAR选系,把它移入较大的盆中,直至每株最后能结2~3个棉铃。这样产生的种子,供大田鉴定和选择之用。
以上程序从F
(三)杂种优势利用
有关主要病害的抗性遗传趋势研究已经肯定,对枯萎病,其F
棉花杂种优势早在1894年,Mell首次描述了陆地棉与海岛棉种间杂交的杂种优势表现。1907年,Balls也报道了陆地棉与埃及棉种间杂种一代,在株高、早熟性、绒长和籽指等性状的优势现象。此后,世界各产棉国对棉花杂种优势利用进行了大量的试验研究。
我国棉花杂种优势研究始于20世纪20年代。冯泽芳等(1923)研究并发现了亚洲棉品种间的杂种优势。奚元龄(1936)研究证明,亚洲棉不同生态型的品种间杂种一代的植株高度、衣指、单铃籽棉重、单铃种子重及纤维长度等性状都表现有显著或微弱的杂种优势。1947年,杜春培等以鸿系265-5与斯字棉2B杂交,开创我国陆地棉品种间杂种优势利用研究之先河。研究结果认为,杂种一代的多数性状有明显的优势,绒长、衣分和单铃重介于双亲之间,生育期偏向早熟亲本。50~60年代的研究工作以陆地棉与海岛棉种间杂种优势利用为主。70年代以后转入陆地棉品种间的
1.亲本选配
大量的研究结果业已表明,不同亲本之间杂交,其F
(1)研究亲子关系
对于利用F
(2)研究亲本间遗传差异
亲本选配是否合理与杂交亲本间的遗传差异有着密切的关系。如何衡量亲本间的遗传差异,通常认为,地理来源差异大的品种反映在形态、生态、生理和发育性状上的差异也大,也许可以用亲本地理上距离的远近代表它们的遗传差异的大小。但进一步研究表明,亲本的地理分布与其遗传差异并无直接联系。由于一个符合育种目标要求的组合(F
(3)配合力分析
配合力是在选育玉米自交系的工作中引出的概念,是指一个自交系与另外的自交系或品种杂交后,杂种一代的产量表现。表现高产的为高配合力,表现低产的为低配合力。目前,配合力的概念已引伸到其他作物的杂
配合力和杂种优势有密切联系,但两者含意并不完全相同。配合力专指杂种一代的经济性状,主要是指产量高低和品质优劣;杂种优势系指杂种在经济性状、生物学性状等方面超越其亲本的现象。因此,利用杂种优势时,既要注意亲本配合力的高低,又要注意它们的杂种在有利性状方面优势的强弱。配合力有一般配合力(GCA)和特殊配合力(SCA)两种。一般配合力是指一个被测系(自交系、不育系、恢复系等)与一个遗传基础复杂的群体品种(系)杂交后,产量与品质等经济性状表现的能力,或这个被测系与许多其他系杂交后,F
(4)外源基因及异常种质的利用
所谓外源基因,指人工构建的或非棉属的其他物种的基因,目前主要是指转
在20世纪90年代以前的20多年中,中国育成的陆地棉品种间杂交棉共13个,当时不存在外源基因参与的可能,也没有异常种质的亲本组成的杂交棉,杂交棉的杂种优势并不十分显著。在20世纪90年代的10年中,育成了23个杂交棉,其中,有转
2.棉花杂种优势利用途径
棉花杂种优势利用途径,实际上就是指杂交制种的方法。尽管杂交制种的方法各不相同,但去雄和授粉是任何杂交制种方法所共有的环节。根据去雄方式的不同,目前,杂种优势利用的途径主要分为人工去雄授粉法、雄性不育系(包括核雄性不育,即一系两用法和核质互作雄性不育,即三系法)、标记性状利用和花器官变异体利用。目前,生产上应用的杂交棉以人工去雄授粉法为主,占95%以上的杂交棉面积,其次是核雄性不育系的利用。
(1)人工去雄授粉法
人工去雄授粉法制种的最大优点就是父母本选配不受限制,配制组合自由,它的缺点是制种全部需要人工操作,工作强度大,种子生产成本高。为了提高制种效率,有关单位做了大量的制种方法研究。湖南澧县研究了冷藏花粉不去雄授粉法,原北京农业大学(现中国农业大学)研究了不去雄强制授粉法,江西省棉花所研究了麦秸套管授粉法,湖北南梓县研究了花冠直套法,河南研究了全株去雄法等,在一定程度上提高了制种效率。但降低制种成本问题仍然没有完全解决。
(2)核雄性不育材料的一系两用法
这种方法又称稳定系自交繁殖制种法。四川省农业科学院经济作物研究所采用这一方法先后选育出的组合有川杂1号、川杂3号、川杂4号、川杂6号等。采用一系两用法,需拔除50%以上的可育株,制种成本仍然较高,其不育系的不育性逐步失去稳定性,达不到1∶1的分离比率;加上不育材料的综合性状已大大落后于现有生产品种,在抗性上难以达到抗病
国内外棉花杂种优势利用均存在制种成本高、优势不很强的共同困难,制种成本居高不下的原因除人工去雄外,各种雄性不育材料的制种环节多也是一个重要原因。核不育材料由于未能解决保持问题,制种过程中必须拔除可育株。核不育两用系仍然要通过系内可育与不育株的授粉来保存。利用核质互作不育材料时,转育恢复基因和育性强化基因周期长,使选拔优势组合大受限制,加大了研制成本。因此,在杂种优势利用上,研究新的技术途径,尤其是生物技术的应用,势在必行。
六、分子育种方法
随着分子生物学、植物病理学及基因工程技术的迅猛发展,运用基因工程手段提高植物的抗病性,为抗病育种开辟了一条崭新途径。分子育种目前主要包括分子标记辅助育种、转抗病基因育种和分子设计育种3部分内容。
分子标记辅助选择(molecular marker-assisted selection,MAS)主要是指基于分子标记和作图技术,利用与目标性状紧密连锁的DNA分子标记对目标性状进行间接选择,以在更短代数内就能够对目标基因的转移进行准确而稳定的选择,聚合多种目标性状特别是对选择隐性基因控制的优良农艺性状十分有利,结合加代技术就可以大大缩短育种年限,缩小育种群体,提高育种效率,聚合选育出抗病、优质、高产的品种。目前,通常采用的分子标记技术主要有RFLP、RAPD、AFLP、SSR等。
国内外一些学者正致力于棉花抗性基因分子标记的研究。一方面通过寻找与抗病基因紧密连锁的分子标记,能够直接或间接地定位抗病基因;另一方面用与抗病基因紧密连锁的分子标记,能够把多个基因聚合在同一个品种中,从而实现抗原积累,提高抗病品种的使用年限,并能把抗性与棉花的其他重要农艺性状结合起来,为分子标记辅助育种提供有力的工具,从而大大缩短育种年限。陈勋基等(2008)以高抗枯萎病的陆地棉品
目前,国内外用于棉花转抗病基因育种的基因主要有两类:一类为病程相关蛋白(PR protein)基因,包括各种来源的几丁质酶基因(
天麻抗真菌蛋白(gastrodia antifungal pratein,简称GAFP)是从我国传统中药天麻(
从7月20日枯萎病发病高峰期调查枯萎病圃抗病性鉴定结果看(表5-9),对照军棉1号的枯萎病发病程度重,枯萎病病指54.59,受
株系发病率(%)病指相对病指抗效抗病类型HB020427.088.858.1483.79RHB020845.4513.6412.5575.01THB024032.919.819.0382.03RHB025412.505.635.1889.69RHB026018.7513.2812.2275.67T军棉1号(CK)86.9954.5950.000.00S新B1(CK)67.9742.8739.4421.47S
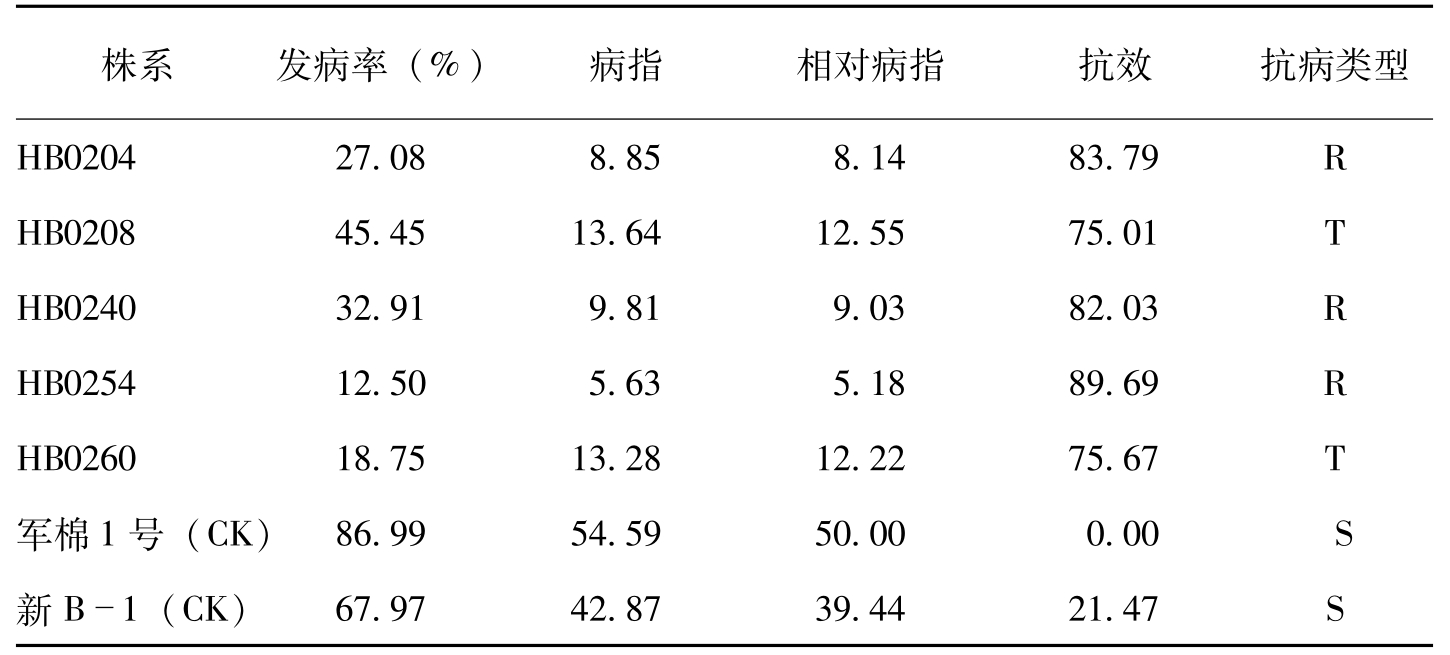
表5-9 转基因陆地棉后代株系抗枯萎病鉴定结果
株系HB0204HB0208HB0240HB0254HB0260枯萎病抗效提高(%)43.0831.9839.0046.6632.64
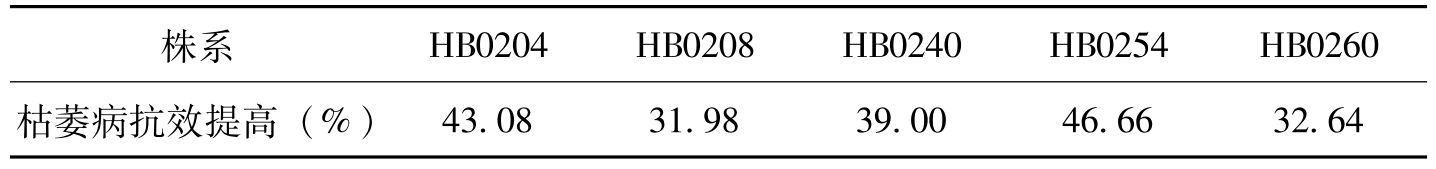
表5-10 转基因陆地棉后代株系与受体品种的抗效比较
编号总株数发病株数(剖秆10月28日)0级1级2级3级4级发病株率(%)病指0305421410400028.67.1030546139400030.87.70305521411300021.45.4030556149500035.78.90305571612400025.06.30305621210200016.74.20306011511400026.76.70306432200000.00.1
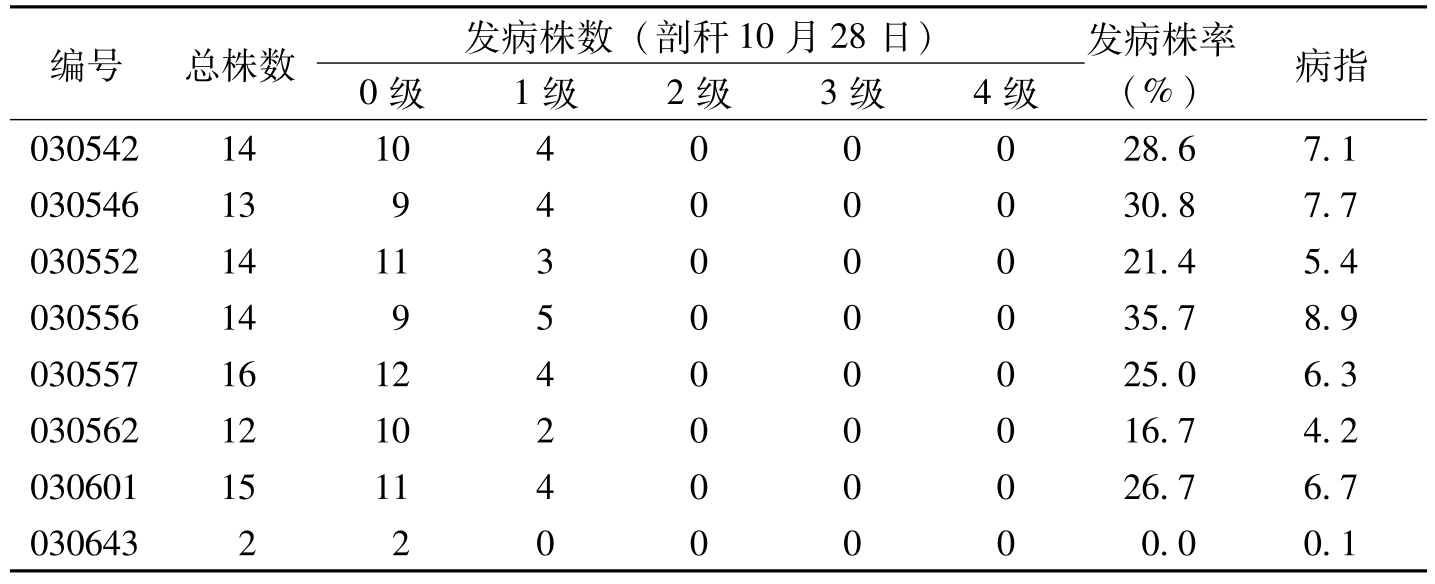
表5-11 2003年枯萎病圃转基因抗病材料发病情况调查(10月28日)
目前,研究较多的是以拟南芥为模式植物的系统获得抗性SAR(system acquired resistance),当植物受到病原菌侵染后局部的HR(hypersensitive resistance)会产生一类信号分子,这种信号分子能诱发整个植株的防卫基因表达,从而使植物对更多的病原菌产生抵制作用(王忠华等,2004)。而
品种总株数第1天第6天第11天第16天第21天病株数发病率(%)病株数发病率(%)病株数发病率(%)病株数发病率(%)病株数发病率(%)转基因军棉1号128001612.52821.93225.04837.5转基因中棉所35153001711.12315.03422.23422.2军棉1号对照150003322.08456.010066.710066.7中棉所35对照150001510.04530.06040.07550.0
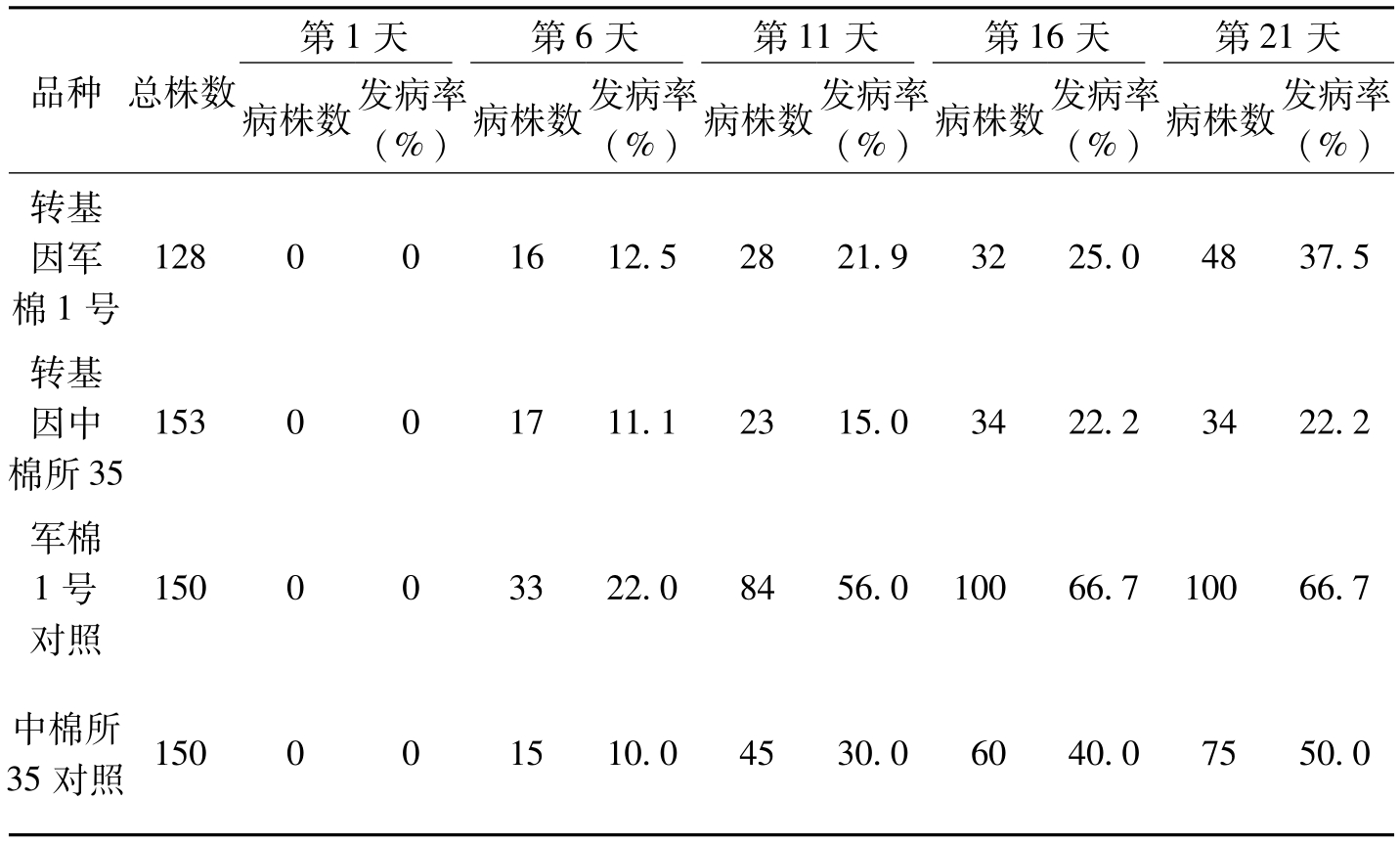
表5-12 接菌后棉株培养钵伤根法接菌发病率统计
利用分子生物学技术揭示抗性提高材料中相对于原感病品种中差异表达的基因,为后续克隆这些抗病相关基因和研究定向筛选抗病性提高的分子机制奠定了良好基础。Liang等(1995)发明的mRNA差异显示(mRNA differential display)技术是一种快速、简便、灵敏的能直接鉴定和克隆差异表达基因的方法。近年来,此方法已被广泛地运用到真核生物基因差异表达的研究中。吕学莲等(2008)以天九63棉花品种和其经过病圃定向筛选2年、3年分别对枯萎病菌7号生理小种表现高感、耐病和抗病的材料为研究对象,利用差异显示PCR技术对接种枯萎病菌前后的抗、感材料中差异表达的基因进行了初步研究,结果共得到62个差异条带。进一步通过反向Northern杂交验证,发现其中有8个是抗病诱导增强的阳性条带。经克隆、测序和同源性比对分析,结果表明,除14-3片段没有找到同源
作物分子设计育种最初由荷兰科学家Peleman(2003)提出,其目的是通过各种技术的集成与整合,在育种家的田间试验之前,对育种程序中的各种因素进行模拟、筛选和优化,确立目标基因型、提出最佳的亲本选配和后代选择策略、提高育种过程中的预见性。分子设计育种一般包括以下步骤:①找到育种目标性状的基因/QTL或其紧密连锁标记;②利用QTL位置、遗传效应、QTL之间的互作、QTL与环境之间的互作等信息,模拟和预测各种可能基因型组合的表现型,从中选择符合特定育种目标的基因型;③进行目标基因型的途径分析,制定育种方案;④根据制定的育种方案进行育种,在此过程中,合理应用分子标记育种、转基因育种和传统育种技术,实现预期目标。由此看来,可把分子设计育种看成分子育种的高级形式。全基因组选择技术,也可认为是分子设计育种的组成部分。
近20年来,随着分子生物学和基因组学等新兴学科的飞速发展,使育种家对基因型进行直接选择成为可能,作物分子育种因此应运而生。分子育种就是把表现型和基因型选择结合起来的一种作物遗传改良理论和方法体系,可实现基因的直接选择和有效聚合,大幅度提高育种效率,缩短育种年限,在提高产量、改善品质、增强抗性等方面已显示出巨大潜力,成为现代作物育种的主要方向。









