棉花枯萎病的生物防治
棉花枯萎病的生物防治
植物病原菌的生物控制是最有前景也是比较实际的一种方法,众多的微生物已被证实具有生防效果(Massarty等,2006)。目前,棉花枯萎病的生防因子比较多,主要有微生物的利用和棉花枯萎病抑菌土的形成。
一、微生物的利用
应用拮抗微生物防治棉花枯萎病是比较有前景的防治方法。目前,用于防治棉花枯萎病的微生物主要包括真菌和细菌等。
(一)拮抗真菌
能够对棉花枯萎病产生拮抗作用的真菌较多,主要是木霉,包括哈茨木霉菌、绿色木霉菌等主要种类。此外,还有青霉菌、黏帚菌、曲霉菌以及非致病性尖孢镰刀菌等,也可作为防治棉花枯萎病的拮抗真菌。
木霉(
1999年,Sivan等对哈茨木霉防治棉花枯萎病的机制进行了研究认为,营养竞争作用是哈茨木霉减少枯萎病菌群体的机制之一。涉及的营养基质有葡萄糖和天门冬酰胺。在非根际土壤中,哈茨木霉不能减少枯萎病菌的数量。Ordentlich等(1991)试验结果表明,在平板对峙试验中,木霉菌
高智谋等(2007)利用平板对峙法和杯碟法测定了从安徽萧县枯萎病田土壤中分离、鉴定获得的哈茨木霉TH-1菌株对棉花枯萎病菌的拮抗作用。结果表明,哈茨木霉TH-1菌株与病菌对峙培养、以及在培养基中加入TH-1菌株孢子悬浮液,对供试棉花枯萎病菌有较好的抑制效果。平板对峙培养2天后,哈茨木霉TH-1菌株和供试病原菌沿接种点连线方向的生长均受到对方的抑制,但前者对后者的抑制作用更为明显。表8-1是培养3天和4天后各供试病原菌单独培养和与TH-1菌株对峙培养沿接种点连线方向的生长距离测定结果。从表8-1可知,TH-1菌株对各供试病菌菌株均有显著的抑制效应,抑制效应大小,菌株间有一定差异。对峙培养4天后,枯萎病菌WWKW1-1、WJKW2-4和XXKW5-1的菌落被哈茨木霉TH-1包围,菌落背面颜色由紫色变为橘黄色,并不再生长,两菌落间形成对抗局面。其后,TH-1可继续向前生长,并部分覆盖于枯萎病菌菌落之上,但最终不能完全覆盖枯萎病菌。说明在对峙培养中,哈茨木霉TH-1菌株的营养和空间竞争能力优于供试病原菌。培养5天后,哈茨木霉TH-1菌株各浓度处理对所有供试菌株的菌丝生长均有显著的抑制作用,且总体来说,抑菌率随TH-1菌株菌液量的增加而呈现递增的趋势。其中,2.50ml的处理对各供试病菌均完全抑制;1.00ml的处理对WWKW1
供试菌株培养3天后生长距离培养4天后生长距离对峙培养(cm)单独培养(cm)抑制率(%)对峙培养(cm)单独培养(cm)抑制率(%)WWKW111.432.6445.831.503.2854.27WJKW241.272.0838.941.302.4346.50XXKW511.432.7147.231.533.2953.50
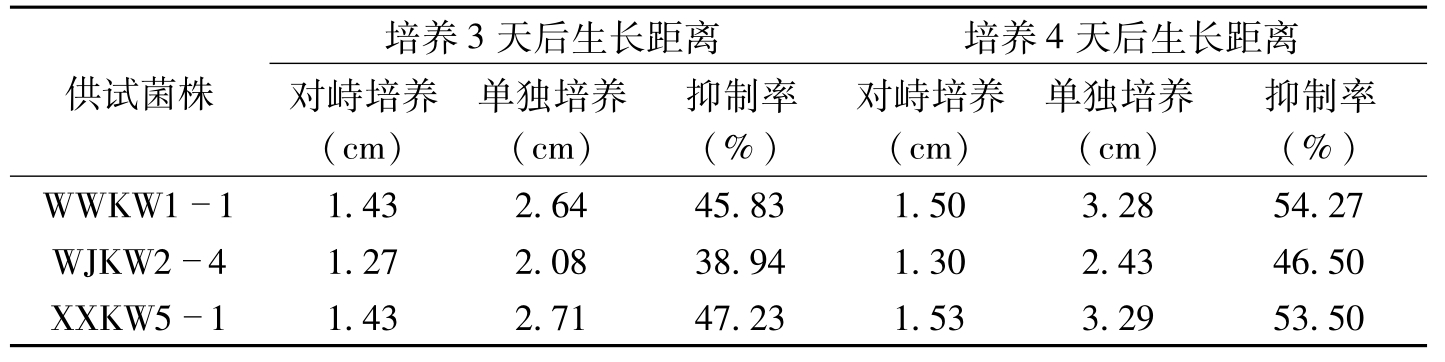
表8-1 平板对峙培养哈茨木霉TH-1菌株对棉花枯萎病菌生长的抑制作用
对棉花枯萎病具拮抗作用的真菌,不仅可从棉田土壤中分离获得,而且也从蘑菇培养料废料和某些昆虫体内分离获得。张军华等(2009)从蘑菇培养料废料中,分离筛选出10株生防真菌。通过室内抑菌试验筛选出生防效果显著的7个菌株:TH、TP、TV、T
编号拮抗菌株R值菌株来源采集地TLTrichodermalongibrachiatum0.6802a棉籽壳平菇培养料东昌府THTrichodermaharzianum0.3980bc棉籽壳平菇培养料东昌府TVT.viride0.4151bc棉籽壳平菇培养料莘县TCT.citrinoviride0.6763a棉籽壳平菇培养料冠县TKT.koningi0.4827b棉籽壳平菇培养料莘县TPT.pseudokoningi0.3827c棉籽壳平菇培养料东昌府T1-1T.sp0.4527b木屑香菇培养料冠县T2-2T.sp0.5927a木屑香菇培养料冠县T1-5Trichodermaspp0.4127bc玉米芯平菇培养料莘县PPeniciliumsp.0.7027a玉米芯平菇培养料莘县CK1—

表8-2 蘑菇培养料分离所得拮抗菌对棉花枯萎病菌抑菌效果
金龟子绿僵菌是一种广谱性的昆虫病原真菌,其寄主范围广泛,能够侵染多种农业害虫,人类利用它防治害虫的历史已愈百年,但国内外有关虫生真菌对棉花枯萎病菌的生防效果鲜见报道。齐永霞等(2010)采用平板对峙培养法和杯碟法测定了金龟子绿僵菌对棉花枯萎病菌的拮抗作用。结果表明,金龟子绿僵菌Ma10、Ma27和Ma55菌株与棉花枯萎病菌WWKW、WJKW和SZKW菌株对峙培养以及在培养基中加入金龟子绿僵菌Ma55菌株的分生孢子悬浮液,对供试棉花枯萎病菌菌丝生长均有较好的抑制作用。平板对峙培养4天后,金龟子绿僵菌菌株和供试棉花枯萎病菌菌株沿接种点连线方向的菌丝生长均受到对方的抑制,但前者对后者的抑制作用更为明显。由图8-1可以看出,绿僵菌Ma55与棉花枯萎病菌WJKW在对峙培养处形成明显的隔离带。液体振荡培养不同时间获得的金龟子绿僵菌Ma55发酵液对棉花枯萎病菌WJKW、WWKW和SZKW的菌丝生长、
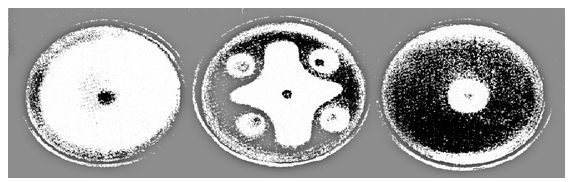
图8-1 金龟子绿僵菌Ma55与棉花枯萎病菌WJKW的对峙培养(4天)
发酵液培养时间(天)抑制率(%)WJKWWWKWSZKW74.20±0.563.96±0.093.35±0.161025.60±0.9223.37±0.6223.38±0.501228.77±0.5226.93±0.5527.10±0.501436.23±0.4534.17±0.6734.54±0.471637.57±0.5136.04±0.4735.89±0.522059.86±1.3856.99±0.6057.09±0.66
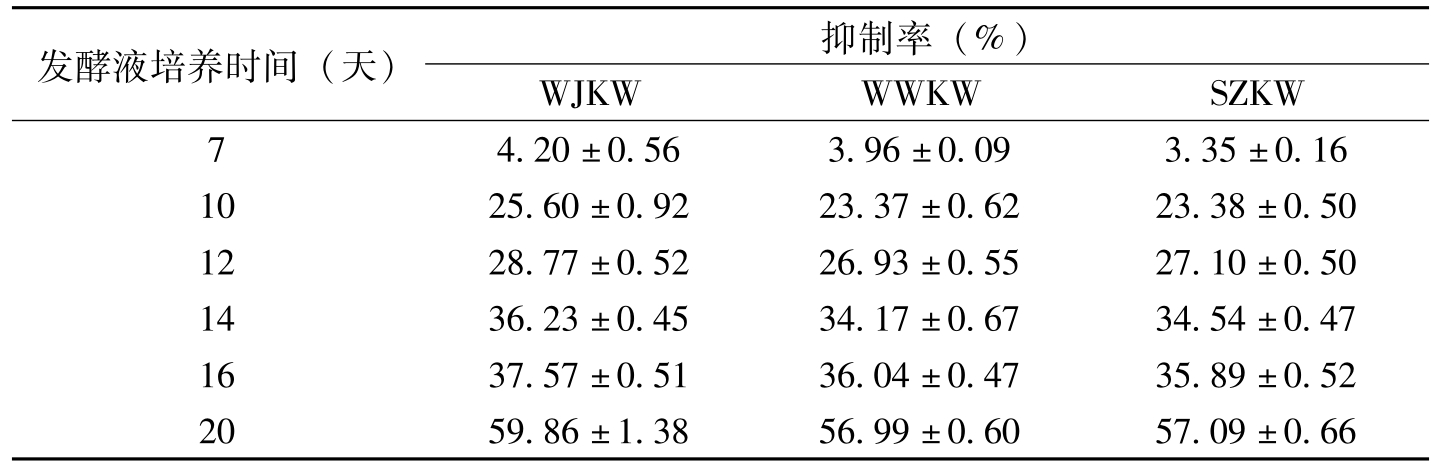
表8-3 金龟子绿僵菌Ma55发酵液对棉花枯萎病菌菌丝生长的影响
发酵液培养时间(天)棉花枯萎病菌分生孢子产生量(×106)(cfu/ml)WJKWWWKWSZKW0(CK)8.90±0.039.13±0.049.21±0.0378.74±0.078.93±0.029.01±0.03107.51±0.047.90±0.028.04±0.02127.03±0.027.28±0.047.51±0.03146.20±0.026.41±0.036.64±0.04165.39±0.025.56±0.045.84±0.04203.14±0.043.37±0.043.91±0.05
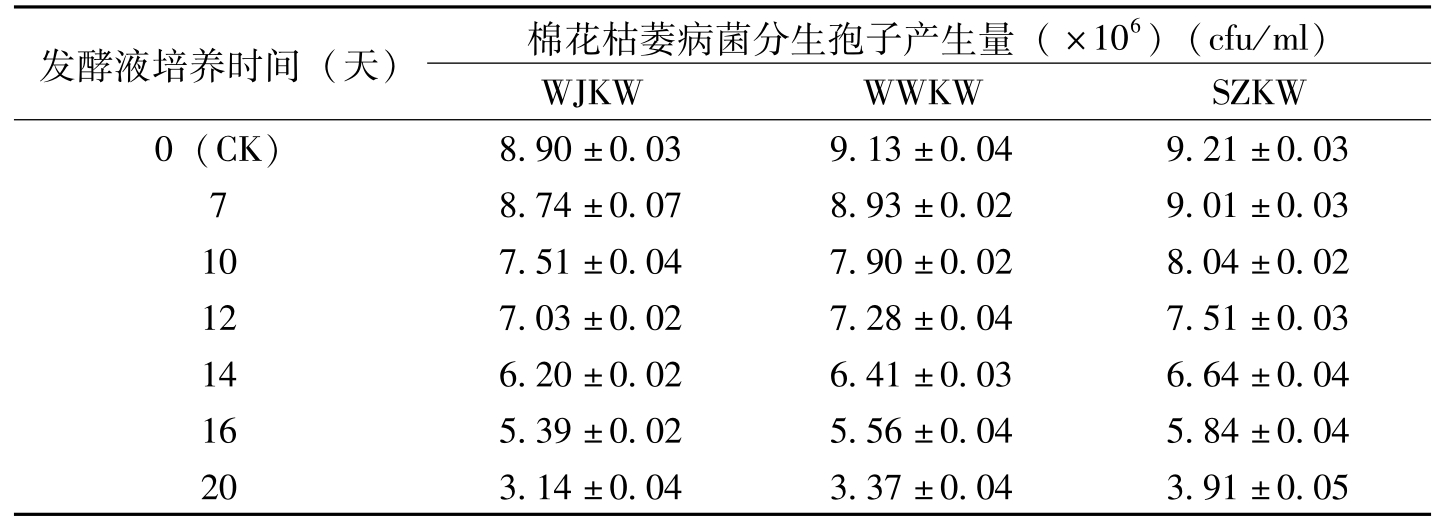
表8-4 金龟子绿僵菌Ma55发酵液对棉花枯萎病菌分生孢子产生量的影响
发酵液培养时间(天)棉花枯萎病菌分生孢子萌发率(%)WJKWWWKWSZKW0(CK)65.89±0.1563.89±0.1162.07±0.94758.94±0.0857.82±0.1858.88±0.461049.83±0.0647.54±0.3648.48±0.501239.94±0.0738.49±0.3039.71±0.571434.54±0.0630.25±0.0633.37±0.671624.21±0.0722.77±0.2023.89±0.262011.66±0.2910.67±0.0913.00±.12
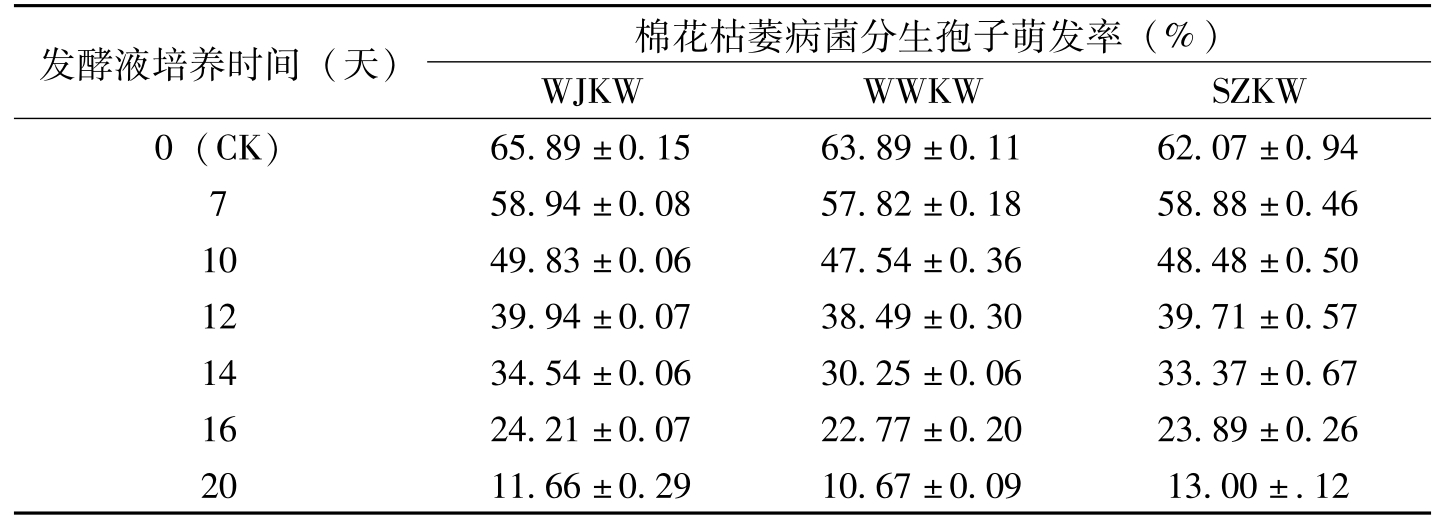
表8-5 金龟子绿僵菌Ma55发酵液对棉花枯萎病菌分生孢子萌发率的影响
绿僵菌素是金龟子绿僵菌分泌的一种次生代谢物质,具有明显的杀虫活性。自从1961年Kodaira在培养金龟子绿僵菌的滤液中首次发现绿僵菌素以来,科学工作者对绿僵菌素开展了广泛的研究,但国内研究较少。1985年,Hino等从绿僵菌属中分离出苦马豆素,发现苦马豆素是一种很有潜力的免疫调节剂。金龟子绿僵菌Ma55发酵液对棉花枯萎病菌的菌丝生长、分生孢子产生量及分生孢子萌发具有较好的抑制作用,与绿僵菌素和苦马豆素的产生是否有关系尚待进一步研究。总之,金龟子绿僵菌是一种重要的昆虫病原真菌。昆虫病原真菌在代谢类型上十分复杂,能产生多种生理功能特异的生物活性物质。这种代谢产物的多样性,为人类开发新的生物防治制剂、药品及在其他领域的利用提供了重要的途径。
来自昆虫的病原真菌球孢白僵菌(
菌株培养3天培养5天生长距离(cm)抑制率(%)生长距离(cm)抑制率(%)038318.54±0.16cdBC20.5519.02±0.16bB44.83445218.05±0.47dC22.6518.55±0.41bB46.29115118.63±0.17cdBC20.1819.11±0.21bB44.58310818.35±0.26cdBC21.3518.79±0.45bB45.39001318.44±0.36cdBC21.0017.94±0.12bB45.14290919.36±0.31bcBC17.0619.33±0.34bB42.41290119.20±0.16bcBC17.7419.33±0.32bB42.88305119.69±0.48bB15.6219.97±0.32bB41.41CK23.34±0.20aA—33.61±3.39aA—
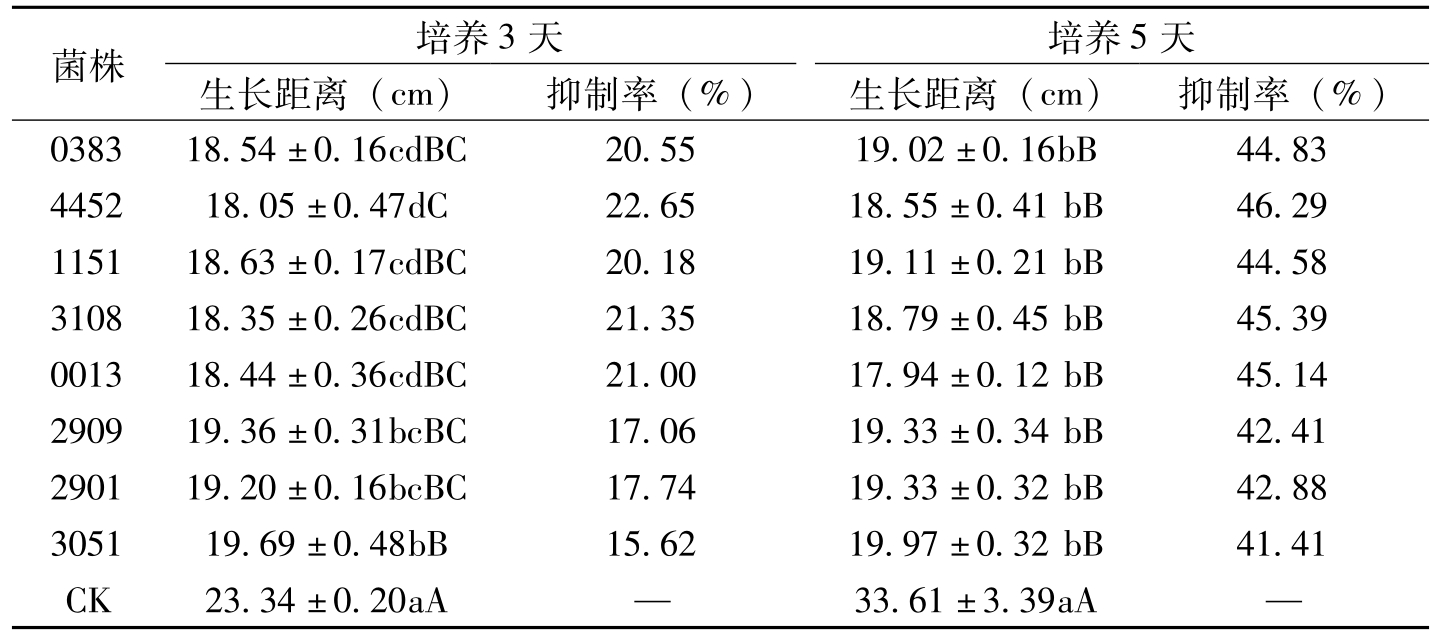
表8-6 8株白僵菌平板对峙培养对棉花枯萎病菌生长的抑制作用
发酵液(ml)3天抑制率(%)5天抑制率(%)2.538.8244.04233.8141.481.534.2035.76123.5728.080.514.1119.83
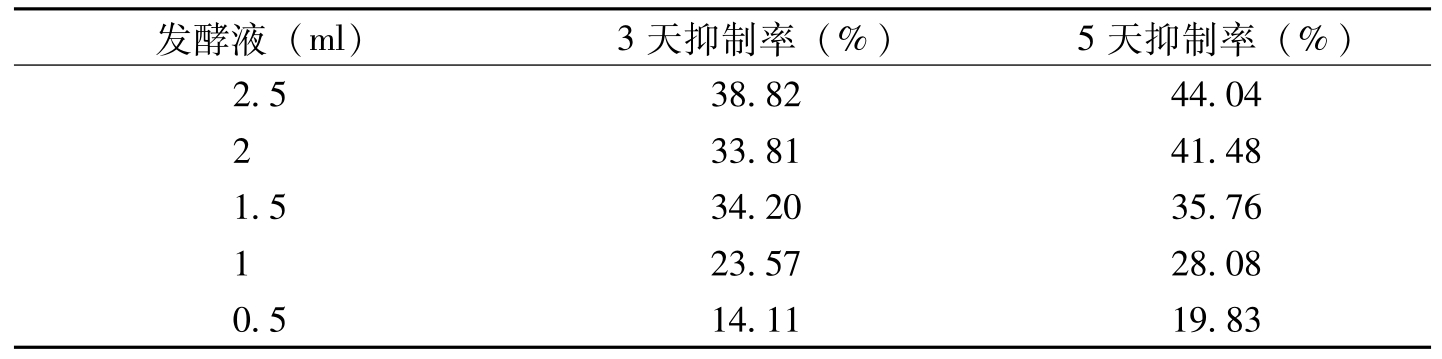
表8-7 白僵菌菌株4452的发酵滤液对棉花枯萎病菌生长的影响
(二)拮抗细菌
目前,已报道的用来防治棉花枯萎病的拮抗细菌主要有假单胞菌[恶
1.内生细菌
内生细菌是指能在健康植物的组织内定殖,在一定的条件下与植物体建立和谐互惠,互相制约关系的一类微生物。实际上50多年以前就已有关于植物体内存在内生细菌而不引起植物病症的报道。人们不断地从植物的根、叶、茎和种子上分离并鉴定出多种植物内生细菌,目前,已从近30多种植物中分离到了近60个属的内生细菌。研究发现,内生细菌寄生在植物的组织内部,其代谢产物不仅对植物本身有各种各样的影响,而且对其他病原生物也有各种不同的作用。内生细菌在植物组织内有足够的碳源、氮源,并且受到植物组织的保护,所以,比暴露于恶劣环境的附生菌和腐生菌更具有稳定的生存环境,易于发挥作用,因此,内生细菌作为生防因子的研究越来越受人们的广泛重视,成为国内外生物防治的又一热点。
棉花植株的内生细菌由许多不同的种群组成。由于棉花各种组织具有不同的生理及营养环境,从而制约着共生的细菌的种类。但一种植物中往往以某一两种细菌为主要种群。鲁素云等(1989)于1979~1984年,先后从北京、河北等7省市棉花枯萎病田采集无病株和轻病株,以及无病田的健株4000余株,其中,包括生产用抗病和感病品种10余个,在我国首次进行了维管中主要微生物类群分析。结果表明,棉株维管中除了定殖引致萎蔫病的病原真菌之外,还有大量的非病原真菌和细菌。维管真菌是以镰刀菌(
属名菌落形态革兰氏染色反应芽孢接触酶氧化酶需氧性测定芽胞杆菌属(Bacilus)圆形或不规则形,灰白,灰色或乳白色直杆状,成对或链状排列+有,椭圆、卵圆、圆形或柱状阳性阳性或阴性好氧或兼性厌氧欧文氏菌属(Erwinia)白色,凸起液状,中心呈稠密凝絮状或呈规则中心环直杆状,单生,成对有时成链—无阳性阴性兼性厌氧假单胞菌属(Pseudomonas)直或微弯中性培养基上蓝色—无阳性阳性或阴性好氧黄单胞菌属(Xanthomonas)黄色直杆菌,多单生—无阳性弱阳或阴性好氧短小杆菌属(Curtobacte-rium)光滑,凸起,常黄色到橙色幼龄培养物中细胞呈小短及不规则杆菌,老培养物变成类球类+无阳性阳性或阴性好氧

表8-8 内生细菌属的主要特征
内生细菌可以随棉花植株的生长而繁殖运转。在棉株生长的各个不同阶段,其中的细菌群体的数量有较大变化,通常内生细菌种群数量从种子开始萌发就开始上升,至花期达最高,此后,则趋于平稳或降低。鲁素芸等(1989)报道,棉花内生细菌数量和组成,随棉株生长发育而变化,从
罗明等(2004)对内生细菌数量测定表明,棉花种子、根、茎、叶柄、叶片等组织内均存在大量的内生细菌。不同品种、组织及种植地,内生细菌的数量不同。各组织中内生细菌的种群密度的分布特点是,种子中最多,其次为根,再次为茎,叶片、花蕾,叶柄中最少(表8-9)。李春宏等(2009)研究了棉花内生菌的数量动态。结果表明:①棉花不同器官的内生细菌数量不同。根中内生细菌数量波动的幅度在lg 4.88~6.79cfu/g-fw,总均值lg 5.59cfu/g-fw;茎和叶波动的幅度在lg 2.597~5.00 cfu/g-fw和lg 2.39~4.60 cfu/g-fw,总均值分别为lg 3.79,3.70cfu/g-fw。根中内生细菌的数量显著高于茎和叶片,这种趋势表现在6个棉花品种取样的不同生育期。②棉花不同生育期的内生细菌数量不同。在根中,苗期内生细菌数量波动的幅度在lg 4.88~5.55 cfu/g-fw,均值lg 5.25 cfu/g-fw;开花期数量波动的幅度在lg 4.72~6.19 cfu/g-fw,均值lg 5.32cfu/g-fw;吐絮期数量波动的幅度在lg 5.24~6.79 cfu/g-fw,均值lg 5.98 cfu/g-fw。除了亚7113开花期的内生细菌数量低于苗期外,棉花根中苗期的内生细菌数量低于开花期与吐絮期。在茎中,苗期内生细菌数量波动的幅度在lg 3.29~4.50 cfu/g-fw,均值lg 3.88 cfu/g-fw;开花期数量波动的幅度在lg 2.97~5.00 cfu/g-fw,均值lg 3.74 cfu/g-fw;吐絮期数量波动的幅度在lg 3.04~
生育期地点品种根茎叶片叶柄蕾苗期鄯善五家渠农大试验田高抗5号7.0×1042.6×103120×104新陆早10号2.4×1030.2×1031.8×103新陆早13号1.0×10410×104150×104草棉15×10410×104金科18号1.5×1041.3×1030.1×103新海18号100×1041.6×1030.2×103石彩6号990×1040.21×108990×104中棉36号210×1043.1×1043.8×103新陆早8号120×1041.4×1037.0×104
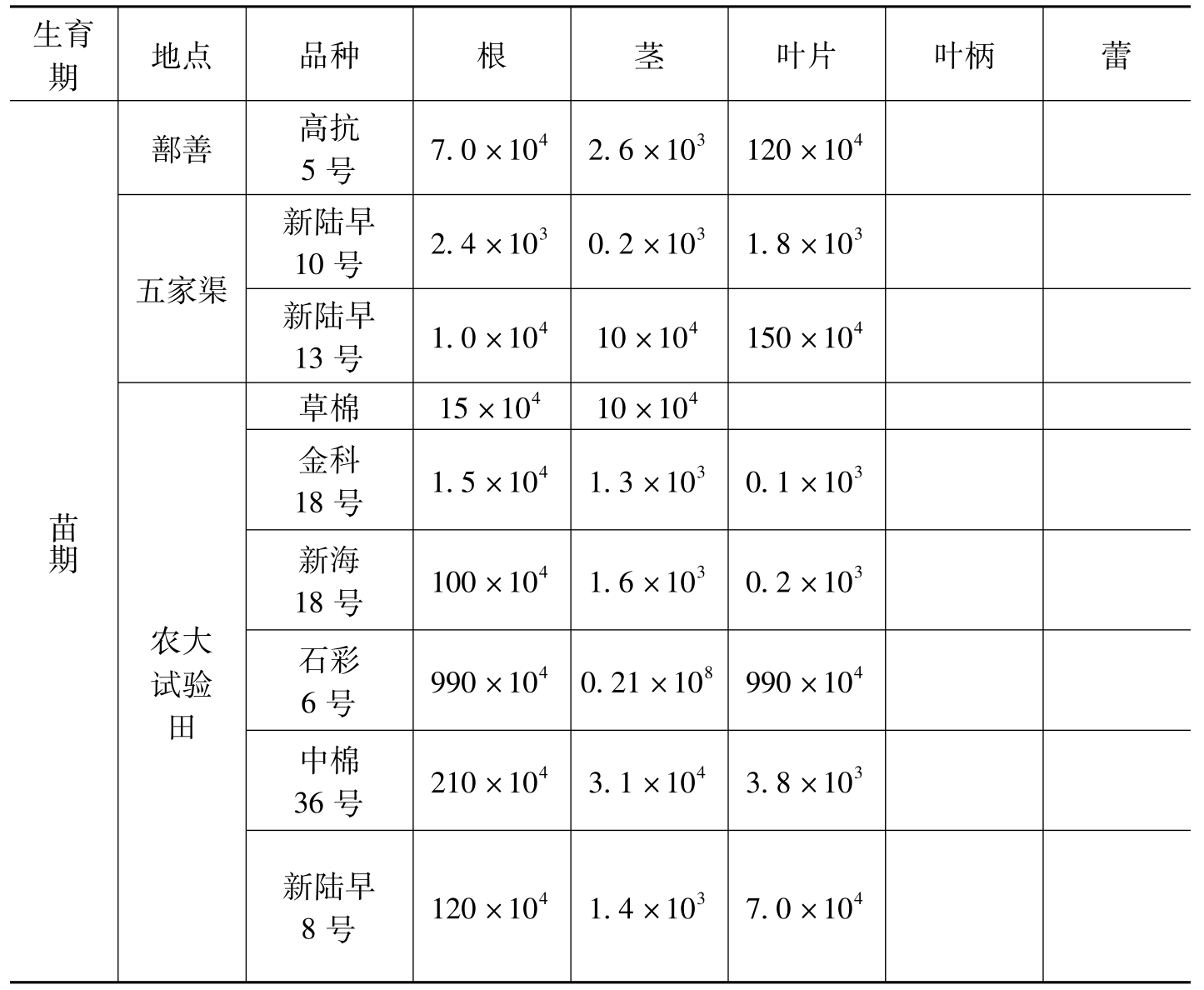
表8-9 不同棉花品种不同时期内生细菌的数量(cfu/g鲜重)
生育期地点品种根茎叶片叶柄蕾温室运早2543.0×1041.2×103180×104新海160.8×1043.0×1030.5×103142100×1046.9×1030.4×104蕾期鄯善农大试验田高抗5号4.0×104300×1043.0×1041.0×1046.9×104草棉3.5×1042.0×1041.4×1041.2×10419×104金科18号1.9×1042.5×1045.1×1039.1×1031.1×104新海18号0.5×1042.0×1042.1×1042.4×10437×104
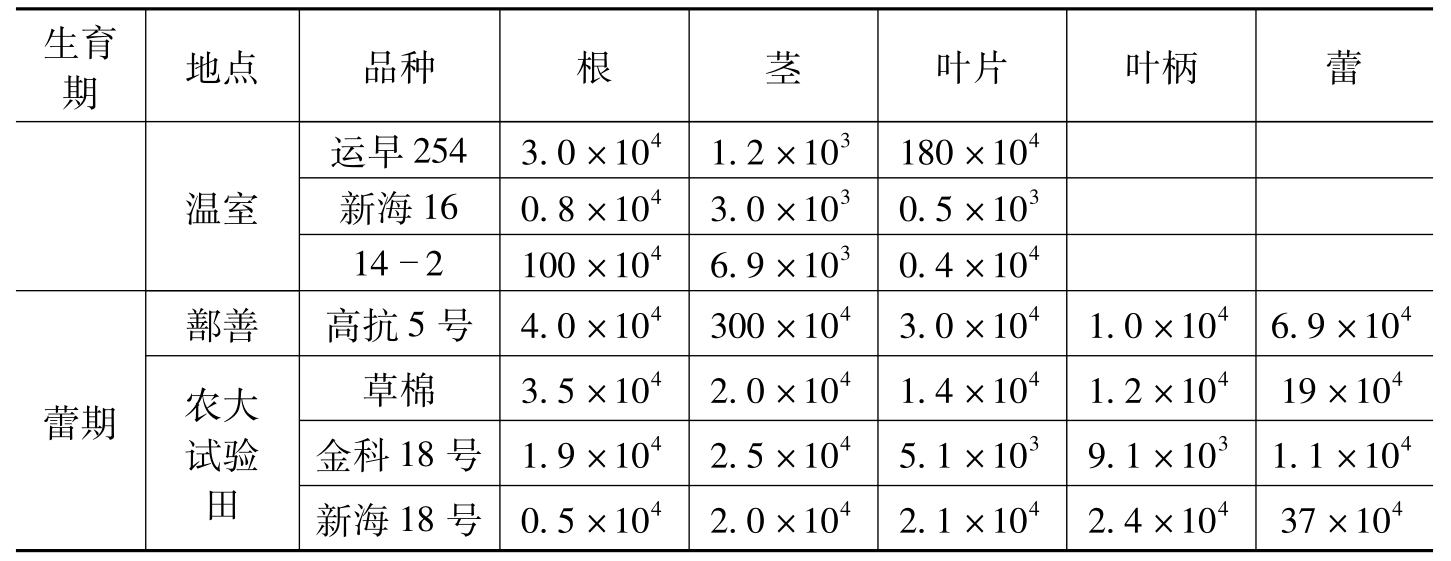
表8-9 不同棉花品种不同时期内生细菌的数量(cfu/g鲜重)(续)-1
内生细菌作为植物微生态系统的组成成分,相对于腐生(附生)细菌、PGPR等生防因子,内生菌更具竞争力,更有利于生防作用的发挥。Chen等(1995)从棉株体内分离到170个细菌菌株,其中,49株已知对棉花枯萎病菌有防治作用,经针刺接种到棉花幼茎上,10天后棉花茎部接种枯萎菌小孢子,12天后枯萎病症状开始出现,利用0~Ⅳ级发病情况计算供试的内生菌对棉花枯萎病的抑制作用。结果显示,其中,6株内生菌在试验中都减轻了棉花枯萎病的发病程度。经鉴定,这6株菌是天牛金杆菌(
菌株抑菌圈半径(mm)菌株抑菌圈半径(mm)0074.00435.00165.70466.00174.00474.50214.50512.00233.70555.00243.50565.50252.50575.00303.50615.80314.00620375.20635.00385.00674.00424.0
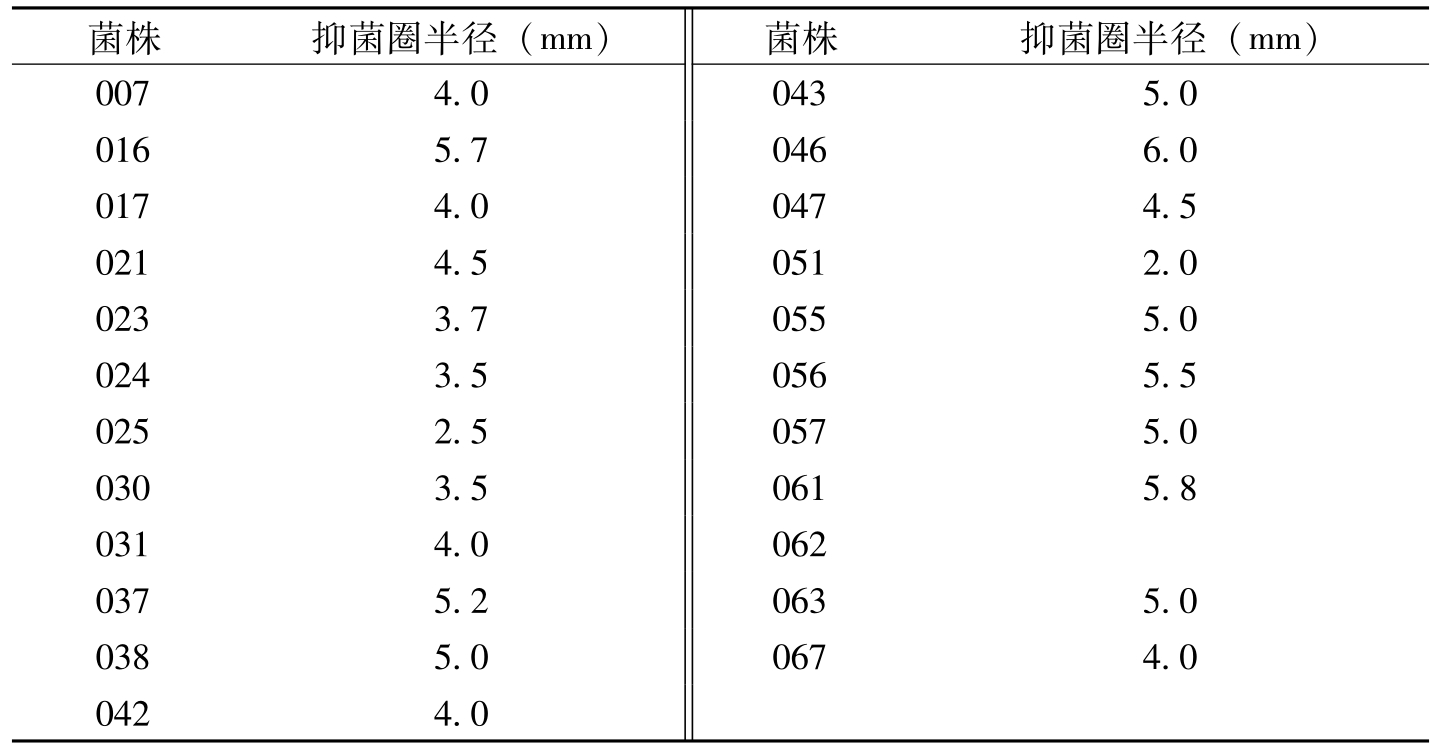
表8-10 内生细菌对枯萎病菌抑菌作用的测定
苦豆子(
通过对550株苦豆子内生细菌进行皿内涂布、对峙培养和胞外分泌物拮抗性试验,筛选到大量的拮抗性苦豆子内生细菌资源。内生拮抗细菌胞
根据菌落特征,10h菌龄的鞭毛染色结果,20h菌龄的菌体形态、大小及革兰氏染色反应,48h菌龄的芽孢染色和72h菌龄的荚膜染色结果,参照东秀珠(2001)的方法进行分类鉴定,具有较强拮抗活性的56株内生细菌分别属于气芽胞杆菌属(
序号分类特征菌株数1气芽孢杆菌属菌体杆状,1.75μm×0.554μm;革兰氏阳性;有芽孢,芽孢囊棒状;侧生鞭毛;无荚膜;菌苔白色,半透明72气单胞菌属菌体杆状,1.83μm×0.73μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔浅黄色,透明8

表8-11 56株苦豆子内生细菌的分类鉴定(龚福明等,2009)
序号分类特征菌株数3芽孢杆菌属菌体杆状,1.51μm×0.55μm;革兰氏阳性;有芽孢,芽孢囊棒状;周生鞭毛;无荚膜;菌苔浅黄色,透明254黄单孢杆菌属菌体杆状,1.53μm×0.43μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔浅黄色,透明55假单孢杆菌属菌体杆状,1.16μm×0.48μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔白色,透明56土壤杆菌属菌体杆状,2.55μm×0.75μm;革兰氏阳性;周生鞭毛;无芽孢;无荚膜;菌苔白色,透明6
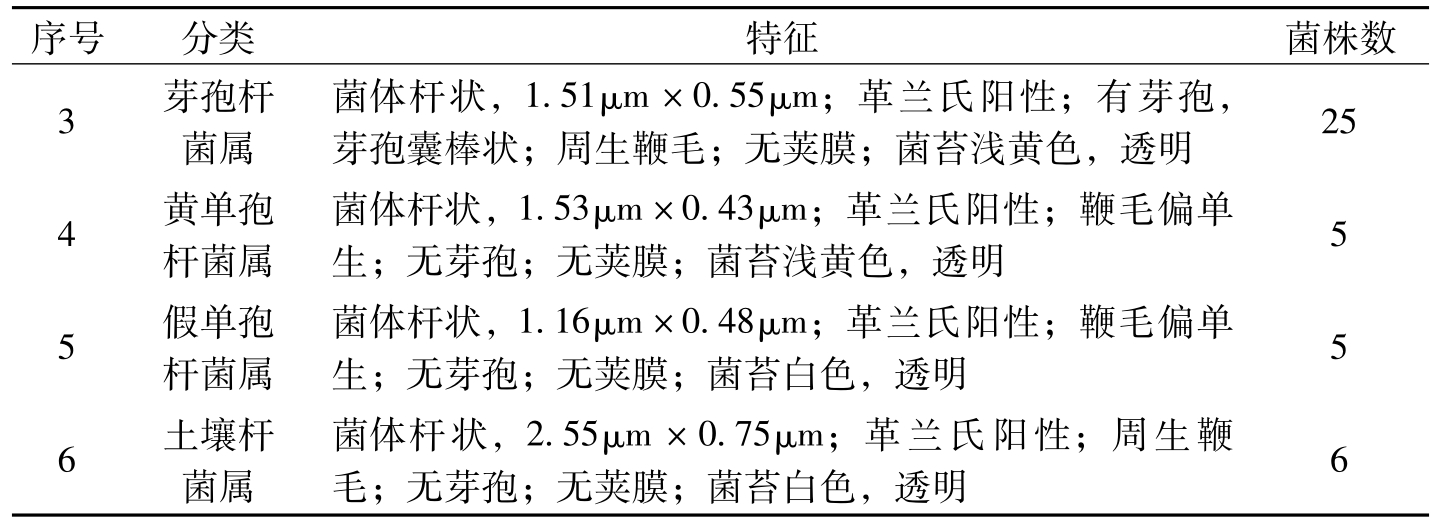
表8-11 56株苦豆子内生细菌的分类鉴定(龚福明等,2009)(续)-1
中国野生甘草分布广泛,主要分布于新疆维吾尔自治区、内蒙古自治区、甘肃、宁夏回族自治区、青海、陕西,河北、西藏自治区也有少量分布。新疆的塔里木河流域是我国甘草蕴藏量、产量最高的地区,有大面积的野生甘草群落。甘草具有很多药用价值,对植物病原真菌也具有一定的拮抗性,甘草内生细菌长期与甘草和谐共存,可能也具有对植物病原真菌的拮抗活性。龚明福等(2007)研究了甘草内生细菌对棉花枯萎病菌的拮抗性,以期筛选出对棉花枯萎病具有生物防治作用的内生细菌,用于生物农药的生产。研究表明,从新疆阿拉尔地区采集野生甘草进行内生细菌分离,共分离得到125份分离物,内生细菌在各个组织部位中的分布是不同的,从根、茎、叶、种子和根瘤中分离得到的内生细菌数分别为35、25、30、15和20。对分离得到的125份内生细菌分离物进行皿内拮抗性试验,结果表明,有31株甘草内生细菌对棉花枯萎病菌表现出明显的拮抗活性,菌落直径1.00~2.86cm,相对抑菌率51.7%~84.7%。这些菌株能否在棉花中定殖,以及在棉花中定殖以后是否仍具有拮抗活性,还有待于进一步研究。根据菌落特征,10h菌龄的鞭毛染色结果,20h菌龄的菌体形态、大小及革兰氏染色反应,48h菌龄的芽孢染色和72h菌龄的荚膜染色结果,参照东秀珠(2001)的方法进行分类鉴定,具有较强拮抗活性的31株内生细菌分别属于气芽胞杆菌属(
序号分类特征菌株数1气芽孢杆菌属菌体杆状,1.75μm×0.554μm;革兰氏阳性;有芽孢,芽孢囊棒状;侧生鞭毛;无荚膜;菌苔白色,半透明22气单胞菌属菌体杆状,1.83μm×0.73μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔浅黄色,透明33芽孢杆菌属菌体杆状,1.51μm×0.55μm;革兰氏阳性;有芽孢,芽孢囊棒状;周生鞭毛;无荚膜;菌苔浅黄色,透明104黄单孢杆菌属菌体杆状,1.53μm×0.43μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔浅黄色,透明55假单胞杆菌属菌体杆状,1.16μm×0.48μm;革兰氏阳性;鞭毛偏单生;无芽孢;无荚膜;菌苔白色,透明56土壤杆菌属菌体杆状,2.55μm×0.75μm;革兰氏阳性;周生鞭毛;无芽孢;无荚膜;菌苔白色,透明6

表8-12 31株苷草内生细菌分类鉴定结果(龚明福等,2007)
包括棉花在内的植物内生细菌种类丰富,开发利用内生细菌作为生物农药的潜力很大。但是关于它对整个生态及人类身体健康影响的报道较少。在防治病害方面,多数生防制剂的防效还不能令人满意,内生细菌的应用潜力还没有完全开发出来。从整体上看,利用内生细菌防治病害存在以下问题:①内生细菌的分离方法尚待改进和完善。内生细菌的分离过程中,灭菌过轻或过重都会影响植物内生细菌调查的准确性,前者会扩大植物内生细菌的生物多样性,而后者会导致内生细菌的丢失;②内生生防细菌的筛选通常是先在实验室的平板对峙培养上进行的,具有很大的局限性,除了拮抗、寄生等作用方式外,许多通过其他方式起作用的菌株不能被筛选到;③内生生防细菌在筛选过程中,往往是采用一种病原菌作为筛选目标,造成生防菌防效单一,综合防病效果差;④内生细菌依赖环境性强,室内试验结果与室外防效不一致;⑤内生细菌作为生防因子,必须考虑其致病性。针对以上问题,必须解决消毒剂的选择与使用问题;在筛选生防菌过程中,采用多种病原菌作为筛选目标,变单一菌剂的使用为多菌配合使用,提高防效和实现防病的广谱性,并降低对环境的依赖性;另外,对土壤中内生细菌的生态学进行更多的研究,使环境和操作更有利于生防细菌作用的发挥;在内生细菌的应用上,必须检测其对人畜和植物的安全性问题。
内生细菌在防治棉花病害的应用上虽然面临许多问题,但作为生防菌株,依然具有广阔的前景。
2.根际细菌
在植物根围区系中存在着大量的微生物,许多微生物及其代谢产物能够抑制植物病原菌的生长发育,因此,从土壤中筛选植物病原菌的拮抗菌,反施于土壤,人为增加有益微生物类群,限制有害微生物生长而进行的生物防治,既可减少化学农药可能带来的环境污染,又解决了抗病棉品种筛选周期长,抗病单一的缺点,被国内外学者广泛研究。
芽胞杆菌(
王少杰(1991)报道了利用对棉花祜萎病菌有拮抗作用的两株荧光假单胞杆菌和两株产芽孢细菌在田间防治棉花枯萎病的试验。结果表明,两株荧光假单胞杆菌的防治效果较好,在苗期的防治效果为73.14%~73.63%,在开花期的防治效果为74.73%~49.73%,棉花增产率分别为15.79%和12.03%。两株产芽孢细菌在苗期和开花期的防治效果分别为62.25%~75.61%和23.98%~55.16%,棉花增产率分别为-0.18%和+4.88%。Zhang(1995)利用枯草芽胞杆菌处理棉种后发现,该菌也能在棉苗根部定殖,并随根的生长而扩展,从而导致枯萎菌在根部的定殖量减少,降低了棉花枯萎病的发生。袁红霞等(1998)报道了两株芽胞杆菌对棉花枯萎病的防治效果为分别36.4%和54.0%。
Gamliel等(1993)利用恶臭假单胞菌(
芽胞杆菌是一类需氧或兼性厌氧、G
芽胞杆菌B
病原真菌920菌株930菌株932菌株大豆根腐病菌+++—+++棉花枯萎病菌+++—+++油菜核盘病菌+++++++小麦赤霉病菌+++—+++水稻恶苗病菌+++—++小麦全蚀病菌++++—小麦根腐病菌+++++++玉米小斑病菌++++++++水稻稻瘟病菌++++++++++水稻纹枯病菌+++—++柑橘黑腐病菌++++++++
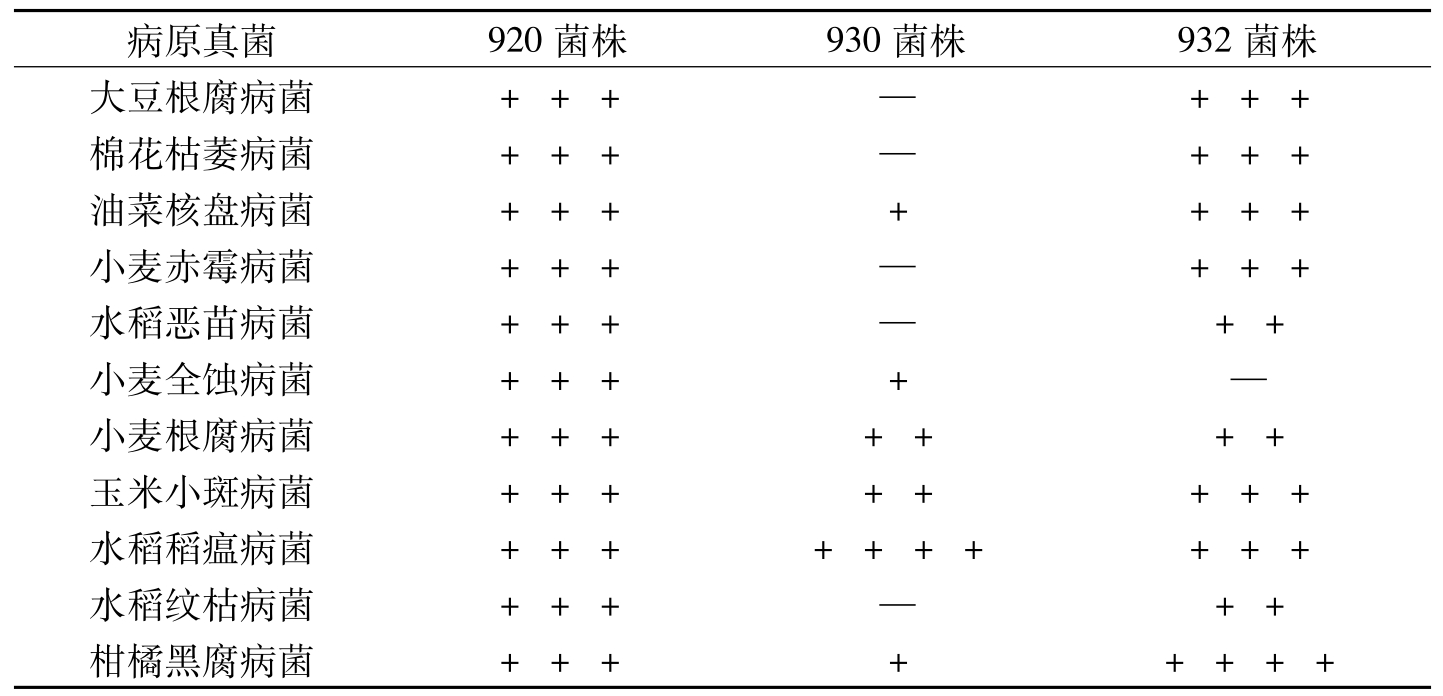
表8-13 3株芽胞杆菌的抗菌谱及抑菌强度测定结果
枯草芽胞杆菌对棉花枯萎病菌的拮抗作用试验结果表明,首先,枯草芽孢菌杆对棉花枯萎病菌菌丝生长有明显的抑制作用。在平板对峙培养中,培养第一天时菌落平均直径为1.1 cm,菌丝向上向外有少许蓬松生长,并且出现明显的抑菌圈,抑菌效果为23.6%。在第三天和第五天时,菌落平均直径分别为1.54 cm和1.46 cm,芽胞杆菌越过抑菌带向菌饼处扩展,菌丝有少许向外生长,气生菌丝消失,菌丝稀薄。抑菌效果分别为54.4%和73.9%。培养至第七天时,菌丝不再向外生长,菌丝非常稀薄,由干燥逐渐变为油状,颜色由白色逐渐变为无色。对照菌丝生长蓬松,菌落厚实,此时已长满皿,抑菌效果为88.8%。其次,枯草芽胞杆菌在培养前期生长速度很快,有很强的占位效应,说明与棉花枯萎病菌存在空间和营养的竞争。显微镜检结果发现,枯萎菌的菌丝相较于正常的菌丝会发生变异,菌丝颜色变褐色、膨大、断裂,消融、释放原生质等物质。说明枯草芽胞杆菌对棉花枯萎病菌的拮抗机制可能还与其分泌的抑菌物质有关,其作用方式和作用机制均有待于进一步研究(但红侠等,2010)。
APS是一种由蜡状芽胞杆菌产生的新型抗真菌环状多肽。刘建国等(1999)研究表明,APS具有广谱抗菌性,其对棉花枯萎病等多种病原真菌及黑曲霉的孢子萌发具有强烈的抑制作用(表8-14),最小的MIC达2.5μg/ml,对真菌的生长亦具有强烈的抑制作用。随着APS处理浓度的增加,菌丝生长速度明显降低,甚至为零。扫描电镜观察表明,经APS处理后,菌丝发生顶端膨大、分支缩短等异常形态学变化。
植物病原真菌处理浓度(μg/ml)菌落直径(mm)抑菌率(%)菌丝生长速度(mm/h)48h72h48h72h48h72h棉花枯萎病菌CK34.543.50.720.60棉花枯萎病菌2514.919.056.955.80.310.26棉花枯萎病菌756.86.880.780.30.140.09棉花枯萎病菌1505.05.085.588.40.100.07小麦赤霉病菌CK33.744.00.700.61小麦赤霉病菌2511.312.066.572.70.240.17小麦赤霉病菌756.06.082.286.40.130.08
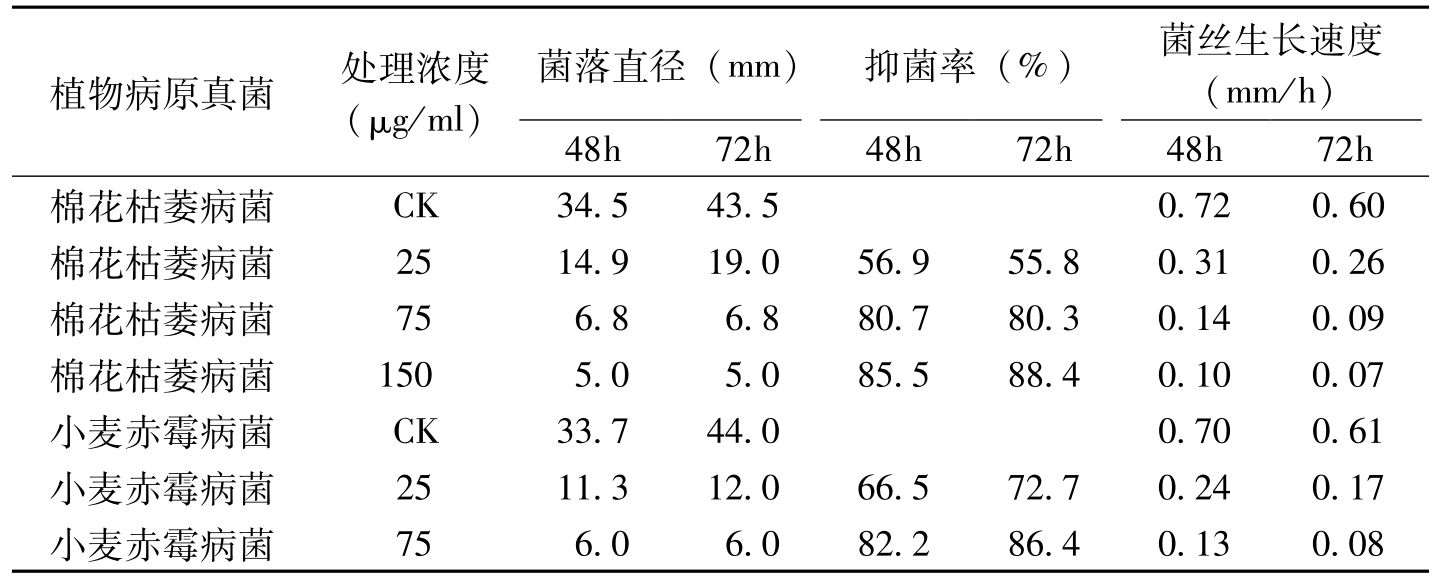
表8-14 APS对病原真菌菌丝生长的影响
植物病原真菌处理浓度(μg/ml)菌落直径(mm)抑菌率(%)菌丝生长速度(mm/h)48h72h48h72h48h72h小麦赤霉病菌1505.55.583.687.50.110.07水稻稻瘟病菌CK32.064.00.670.89水稻稻瘟病菌2510.512.068.781.40.220.14水稻稻瘟病菌756.06.081.290.60.130.08水稻稻瘟病菌1506.06.081.290.60.130.08
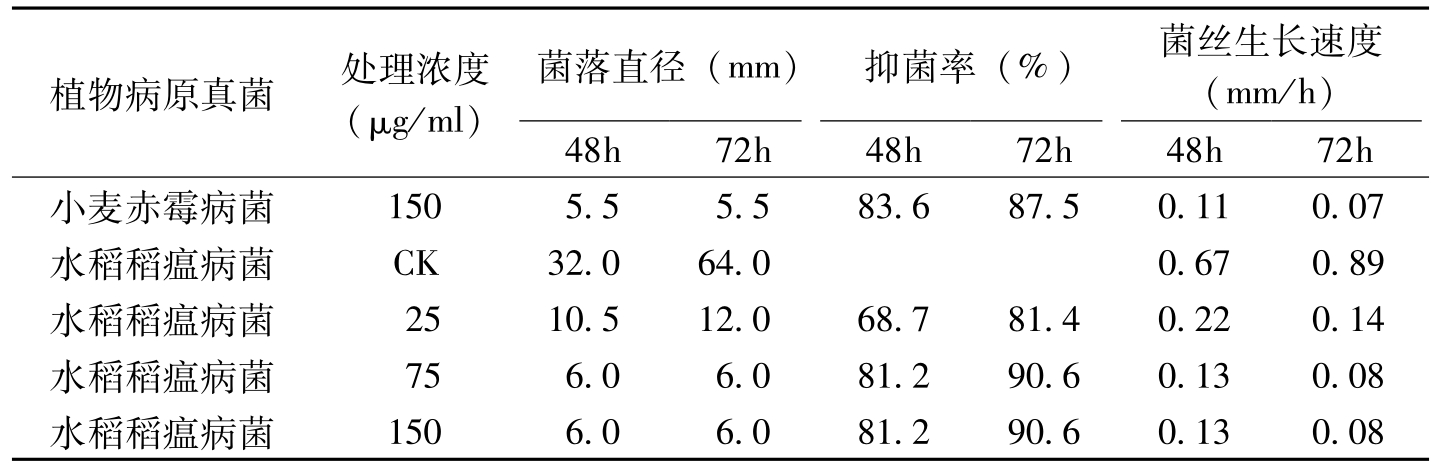
表8-14 APS对病原真菌菌丝生长的影响(续)-1
关于枯草芽胞杆菌对植物病原菌的作用方式,有人认为,是病菌孢子萌发受到抑制,也有报道是菌丝顶端的畸形。孔建等(1998)将枯草芽胞杆菌B-903菌株液体培养72h后,将培养滤液高温灭菌,以不同比例将培养滤液加入镰刀菌孢子悬浮液中,置于扫描电镜和光学显微镜下定时观察。结果表明,镰刀菌孢子悬浮液培养12h后,无菌水处理的病菌孢子均已萌发形成细长菌丝,培养至18h菌丝生长正常,长而均匀,有分枝,培养至48h菌丝未见异常。但经B-903滤液处理的镰刀菌,培养12h后在光学显微镜下观察,孢子尚有部分未萌动,孢子短粗变形,已萌动的芽管出现扭曲状,但多数孢子随后仍能发育成菌丝。上述异常现象随B-903滤液处理浓度增高而加剧。将镰刀菌孢子悬浮液培养15h在扫描电镜下观察,经B-903滤液处理的菌丝端部和中部的细胞开始明显膨胀呈球状,球状细胞的细胞壁皱缩干瘪,分析应是内含物外泄后所致。小孢子端部亦膨大形成远大于自身的球状结构,并看到内含物外泄的现象。经B-903滤液处理18h在光学显微镜下看到,所有的菌丝细胞均出现畸形。细胞呈圆形或椭圆形,以致整个菌丝由丝状变为捻珠状,此时透过细胞壁,其细胞内含物清晰可见。然而至28h,球状细胞进一步膨大,胞内物质减少,细胞透明度增加,并且菌丝开始有断裂现象。36h后观察,串珠状菌丝已纷纷断裂成片段,5~7个球体细胞连成一串,细胞内含物消失,变成空胞,镜下看去类似一串串的肥皂泡。至48h,细胞之间相互离解,菌丝体完全崩溃,最终这些单个的圆形空胞也逐渐瓦解消融。试验中观察到,不同浓度的B-903滤液处理,病菌孢子和菌丝的畸变过程是类似的,而高浓度的
利用木霉菌属、芽孢菌属和假单胞杆菌作为生防制剂防治棉花病害已经取得进展。张克诚等(2002)利用一株从小麦根际筛选的链霉菌属(
二、抑菌土的形成
植物病害抑菌土(Pathogen suppressive soil)是指在土壤中有某种病原菌存在下,种植感病植物,由该病原菌引起的病害的发生、发展受到抑制的土壤。抑菌土的形成须具备两个条件:一是土壤中存在足够以致病的病原菌的量;二是同时种植感病品种,而植株不发病,或发病很少。
从抑菌土的发现到21世纪初,对抑菌土的研究已有100多年的历史。马存等(2007)将抑菌土的研究历程划分为4个阶段。第一阶段:从19世纪末至20世纪20年代,主要是对不同类型土壤病害发生情况的研究报道,是抑菌土研究的初创时期。研究结果只是相继证实了这种现象,均未涉及土壤中是否含有致病菌,以及病原菌含量上的差别。第二阶段:从20年代末到50年代末,Menzies(1959)提出抑菌土概念为止。抑菌土的概念已初步显现。第三阶段:从60年代起至70年代末,由于抑菌土概念的提出以及对环境保护的重视,使植物病理学家对非化学方法保护农作物抵御各种病害的侵袭日益重视,使抑菌土的抑菌机理研究逐步深入,以期从中发现病害防治的新途径。第四阶段:从70年代开始至今,抑菌土的本质得到初步揭示,对其机制的研究逐渐从单一集中于生物因子的主导作用,发展为生物因子和非生物因子共同作用的结果。对抑菌土的研究已成为植物病害生物防治的一项重要内容。
棉花枯萎病重病田或枯萎病圃,连续种植抗病品种多年后,再种植感病品种则发病显著降低,称做病圃衰退。徐富有(1985)认为,抗枯萎病品种连作3年后土壤内枯萎菌致病力显著减弱。张卓敏等(1980)和吴传德(1985)认为,土壤内枯萎菌量随抗病品种连作年限增加而减少,致病力也有随连作年限增加而降低的趋势。中国农业科学院植物保护研究所河南新乡县王屯基点,1972年在发病率接近100%,死苗率80%的枯萎病绝产田块建立枯萎病圃,1973年开始种植抗病品种,到1976年再种植感病品种岱字棉15、发病率降低到35.4%,病指22.3,该病圃在1982年、
(一)抑菌土的抑菌效果
1.盆栽试验
马存等(1992)于1987~1988年两年进行盆栽试验结果表明,86-1根围土(抗病品种86-1连作10年以上田块的棉株5cm内土壤)枯萎病平均发病率37.3%,病指19.5,与对照(无植棉史的果园土或种植1~2年感病棉花品种的土壤)相比,抑菌效果达60.5%。86-1田间土(抗病品种86-1连作10年以上田块土壤)枯萎病平均发病率48.3%,病指29.4,抑菌效果39.9%。枯萎病圃土(作为棉花育种抗病性鉴定病圃连作棉花10年以上病圃土壤)平均发病率55.3%,病指27.2,抑菌效果43.9%。感病品种棉田土发病率72.8%,病指66.9,无抑菌效果。对照果园土平均发病率70.1%,病指48.7(表8-15)。以上结果表明,86-1根围土、86-1田间土、枯萎病圃土对枯萎病菌均有显著的抑菌效果,86-1根围土抑菌效果最强,比病圃土效果高27.4%。感病品种棉田土和果园土对枯萎病菌无抑菌效果。
土样发病率(%)病指抑菌效果(%)861根围土37.319.560.5861田间土48.329.439.9枯萎病圃55.327.243.9感病品种田72.866.9—果园土(CK)70.148.7—861根围灭菌72.455.6—861根围+果园(1∶1)55.237.324.1
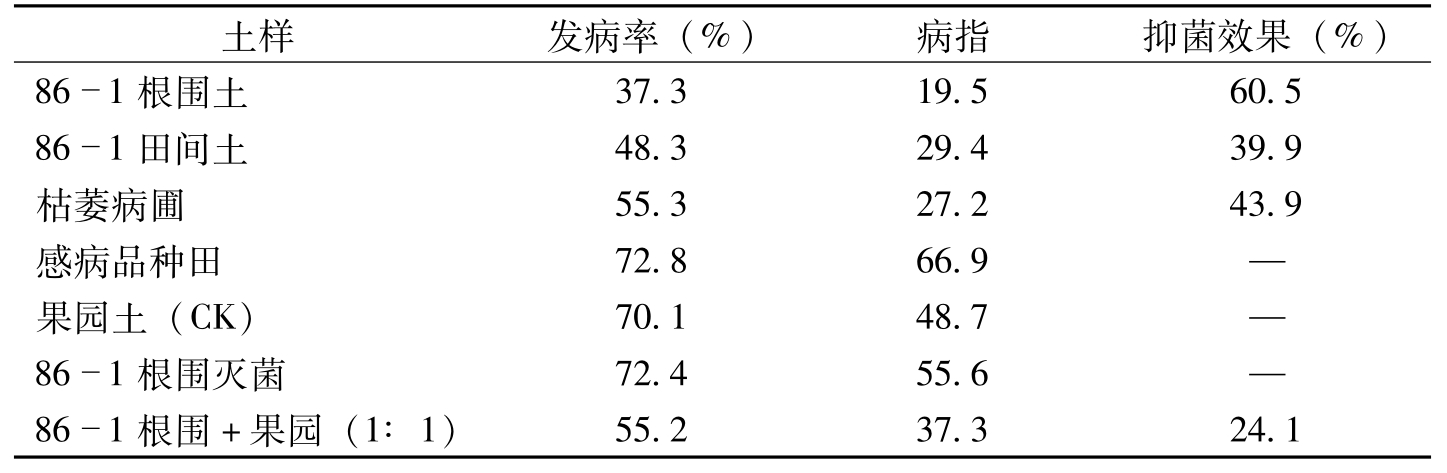
表8-15 不同土样对枯萎病菌抑菌效果比较(盆栽)
李长兴等(1996)报道的盆栽试验结果指出,种植抗病品种10年以上的重枯萎病圃再种感病品种(辽棉6号),发病率为80.2%,病指为
处理项目枯萎病发病率(%)病指抑菌效果(%)抑菌土(10年)80.247.826.5抑菌土(5年)93.258.69.9抑菌土(高压灭菌)86.158.210.4小麦田土(CK)92.165.0—

表8-16 盆栽试验的抑菌效果
2.田间小区试验
河南新乡田间小区试验结果,86-1连作10年田枯萎病平均发病率9.8%,病指4.4,与对照果园田比较,抑菌效果达85.8%。连作10年以上枯萎病圃,平均发病率17.1%,病指6.8,抑菌效果75.2%。对照果园田平均发病率57.l%,病指28.8(表8-17)。在辽宁辽阳试验结果,连作10年枯萎病圃枯萎病发病率39.0%,病指19.3,对照菜园土发病率75.9%,病指44.1,抑菌效果平均56.2%(表8-18);连作5年的枯萎病发病率66.7%,病指37.8,抑菌效果17.6%。发病率和病指比连作10年病圃分别高27.7%和18.5%,而抑菌效果低36.6%。还可看到未接菌小区连作10年和5年病圃田,枯萎病指分别为8.1和14.1,这说明病圃连作年限愈长,病圃衰退愈明显,而对枯萎病菌抑菌效果也就愈显著。河南新乡、辽宁辽阳两地田间小区试验结果基本一致均证明,连续种植抗病品种多年后的棉田,或连作多年的枯萎病圃,对枯萎病菌均有抑制作用,存在抑菌土。
田块处理枯萎病发病率(%)病指抑菌效果(%)861连作田接菌9.84.485.8未接1.00.3—枯萎病圃田接菌17.16.8—未接1.90.676.2果园田(CK)接菌57.128.8—未接0.00.0—
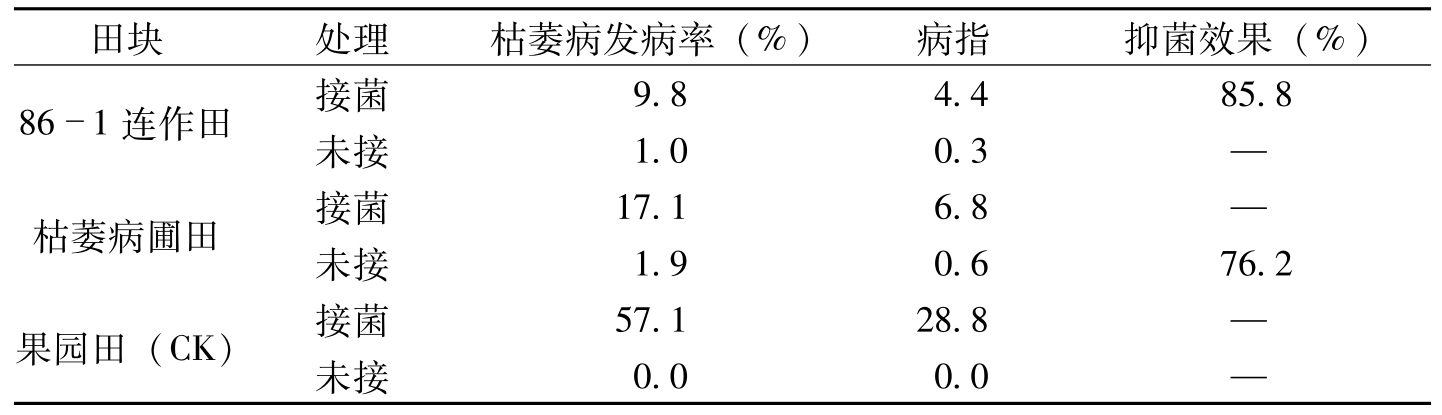
表8-17 田间小区对枯萎病菌抑菌效果比较(河南新乡)(马存等,1992)
田块接菌情况发病率(%)病指抑菌效果(%)连作10年病圃田接菌39.019.356.2不接菌15.58.1—果园田(CK)接菌76.944.1—不接菌0.00.0—连作5年病圃田接菌66.737.817.6不接菌28.514.1—果园田(CK)接菌71.045.9—不接菌0.00.0—
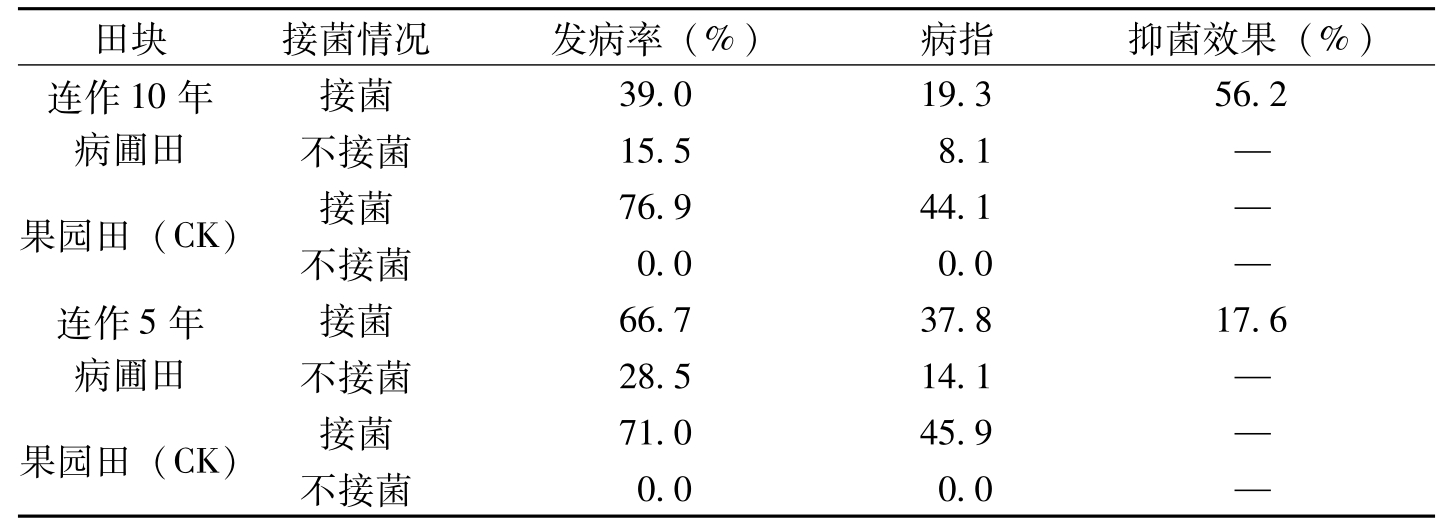
表8-18 不同田间小区对枯萎病菌抑菌效果比较(辽宁辽阳)(马存等,1992)
杨之为等(1995)按不同年限的土样分别接入1%土样的棉枯萎病菌麦粒沙培养物,当种植感病品种冀棉11时,从苗期病情看,连作3年、6年、8年、9年、11年和15年土样的发病率依次为66.67%、55.6%、42.8%、33.3%、11.7%和3.3%,松林土(CK)发病率为85.7%(表8-19)。抗病品种岱字棉16连作6年的土样发病率为CK的64.8%,连作11年、15年的土样发病率仅为CK的13.6%和3.8%。将高压灭菌土样再接入等量病菌培养物,测定其土样致病力,可以看出(表8-19),当抗病品种连作少于8年时,土壤中抑病因子大多被降解,棉苗发病率81%~92%;但连作9年以上则发病率在33.3%~63.3%,低于重病田,说明经高压灭菌的土样中仍可能存在少量抑病因素。
土样来源原土样灭菌土样发病率(%)病指发病率(%)病指松林土85.771.466.354.5重病田土40.020.085.771.4连作3年66.733.381.863.6连作6年55.630.6——连作8年42.817.691.668.7连作9年33.330.533.316.7连作11年11.75.963.650.0连作15年3.30.0156.640.0
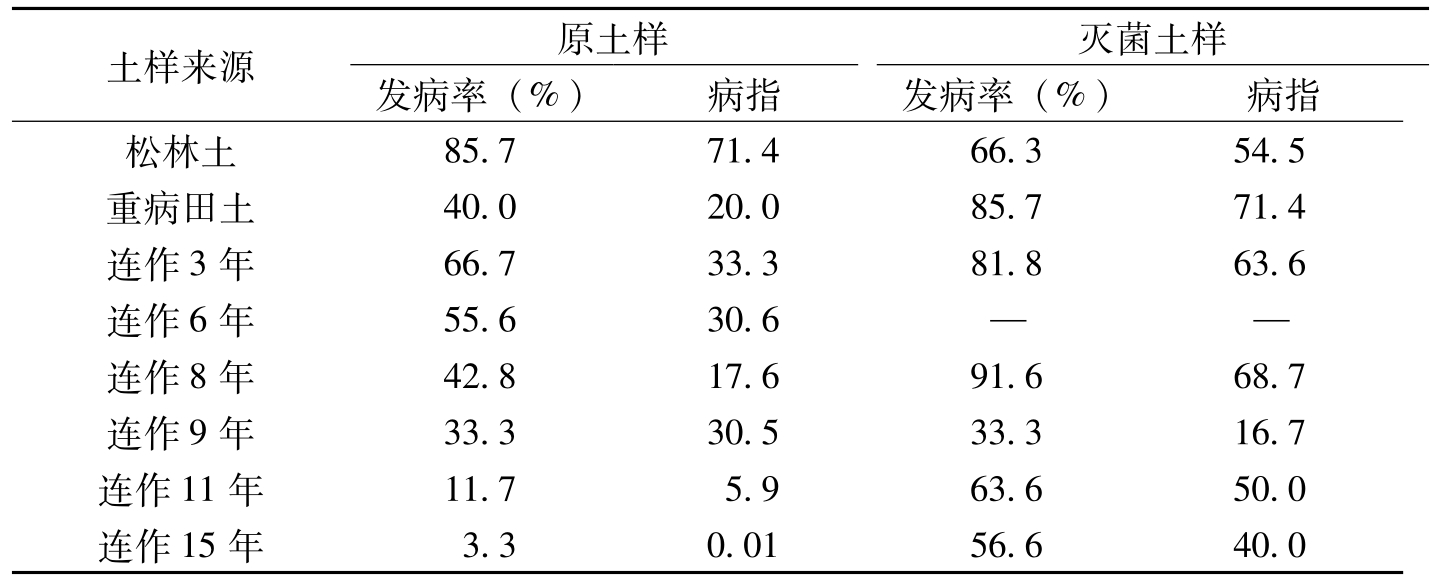
表8-19 不同土样接菌后的抑病效果(接菌量1%)
(二)形成抑菌土的生物因子
形成抑菌土的因子主要有生物因子和非生物因子,不同抑菌土起作用的因子各有不同。在生物因子中,主要是土壤中的微生物。非生物因子是指土壤的矿物质含量、土壤的酸碱性、土壤质地、土壤颗粒结构等物理和化学性质。
国际上有研究认为,土壤微生物种群结构的改变,尤其是一些拮抗微生物的增加,是形成棉花枯萎病田抑菌土的主要生物因子。杨之为等(1995)和王汝贤等(1998)从棉花根系分泌物对棉花枯萎病的影响和连作抗枯萎病品种棉田微生物数量变化做了系统研究后认为,棉花抗病品种连作田土壤中,棉枯萎菌数量的减少,是土壤致病力下降的主要原因,而造成棉枯萎菌数量减少的主要原因,则是土壤中的有益放线菌、细菌增长导致棉花抗病品种根系分泌物中的抑菌物质对病菌抑制的结果。土壤有益微生物增长的原因:一是棉花抗病品种根系分泌物为这些有益菌提供了良好的生活条件;二是可能因棉花枯萎菌数量的减少,有益菌减少了竞争对手,使其有了扩大繁殖的机会,这一切变化又均与棉花抗病品种连作分不开。
1.抗病品种连作不同年限棉田带菌量和致病力
抗病品种连作3年,土壤带菌量约为重病田土样菌量的43%,连作6年为重病田土样菌量的57%;连作8年的土样带菌量降低幅度较大,为重病田的23%;连作10年以上的土壤中,棉枯萎病菌很少(表8-20)。在不同连作年限的抑菌土中种植感病品种冀棉11,重病田的棉苗发病率为31.25%;抗病品种连作3年的棉苗发病率为12.5%;连作6年的发病率为9.09%;连作9年以上的除连作11年发病率4.55%外,其他基本不发病。这表明,随着抗病品种连作年数的增长,土壤中,棉枯萎病菌的数量和致病性逐年降低,说明棉枯萎病抑菌土的形成是抑病因子逐渐积累的过程。
土样来源土样带菌量(个/克土)发病率(%)病指松林土(CK)000重病田土217631.7514.06连作3年114712.509.38连作6年12369.094.56连作8年5008.304.00连作9年5800连作10年1000连作11年1484.564.56连作15年000
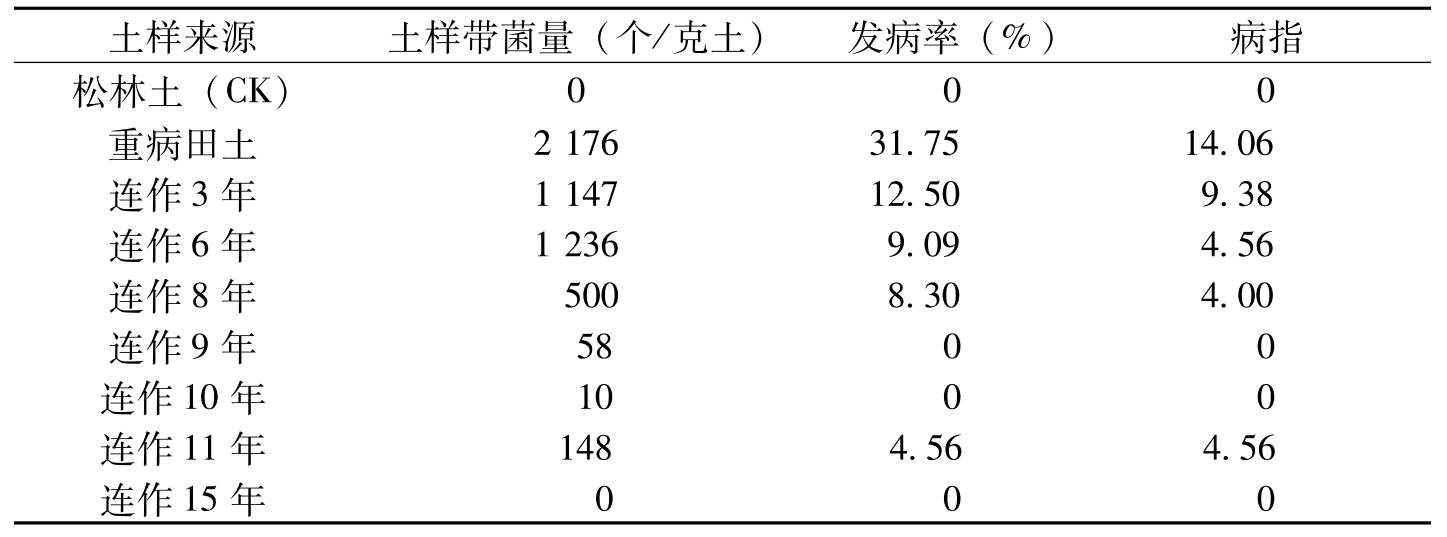
表8-20 不同棉田土样致病力比较(杨之为等,1995)
马存等(1992)在盆栽土样内枯萎菌菌量测定结果表明,86-1根围土,每皿平均有菌落22.5个(每克土样内有菌落4500个),比对照果园土减少56.7%。86-1田间土每皿有菌落21.4个(每克土有4250个),比果园土减少59.2%。枯萎病圃土每皿有菌落32.2个(每克土有6440个),比果园土减少52.9%。对照果园土每皿有菌落52个(每克土有10400个)。田间小区内枯萎菌菌落,86-1田间土、枯萎病圃土每皿有菌落分别为138个(每克土有2760个)和1486个(每克土有2920个),分别比菜园土减少51.4%和48.6%。菜园土每皿有菌落284个(每克土有5580个)。从上述结果清楚的看到,在相同接菌量情况下,种植一季感病棉花品种后,3个抑菌土土样内,枯萎病菌菌量比对照菜园土减少48.6%~59.2%,说明抑菌土对接入土壤内的枯萎病菌有极显著的抑制作用,因而枯萎病发病率显著降低,达到控制枯萎病为害的效果。
2.抗病品种连作棉田微生物数量的变化
王汝贤等(1998)从棉花抗病品种连作3年、6年、8年、9年、10年的土样中分离出3830个真菌菌落,其主要类群归入11个真菌属。随着棉花抗病品种连作年限的增长,土壤中,真菌和放线菌的种类和数量逐渐增加(表8-21、表8-22)。对棉枯萎病菌抑制性较强的种类有:细黄链霉菌(
菌类连作年限10年9年8年6年3年菌落数百分率(%)菌落数百分率(%)菌落数百分率(%)菌落数百分率(%)菌落数百分率(%)脊霉菌属9.448.48.455.36.550.010.163.99.659.3曲霉菌属6.131.43.523.03.829.23.220.34.729.0链格孢属0.84.10.74.60.64.60.31.90.53.1毛壳属0.21.00.95.90.32.31.59.50.21.2园酵母属0.31.50.10.70.32.30000黑粘座孢霉属0.10.5000.21.50.10.60.10.5瓶粳脊霉属00000.10.80.21.300毛霉属000.10.70.21.50.10.600木霉属00000.10.8000.21.2粘帚霉属000.10.70.21.50000镰孢属0.52.60.32.00.21.50.10.60.31.9其他2.010.51.27.20.54.00.31.70.83.8平均每皿菌落总数19.415.313.015.916.2种类数1812141110
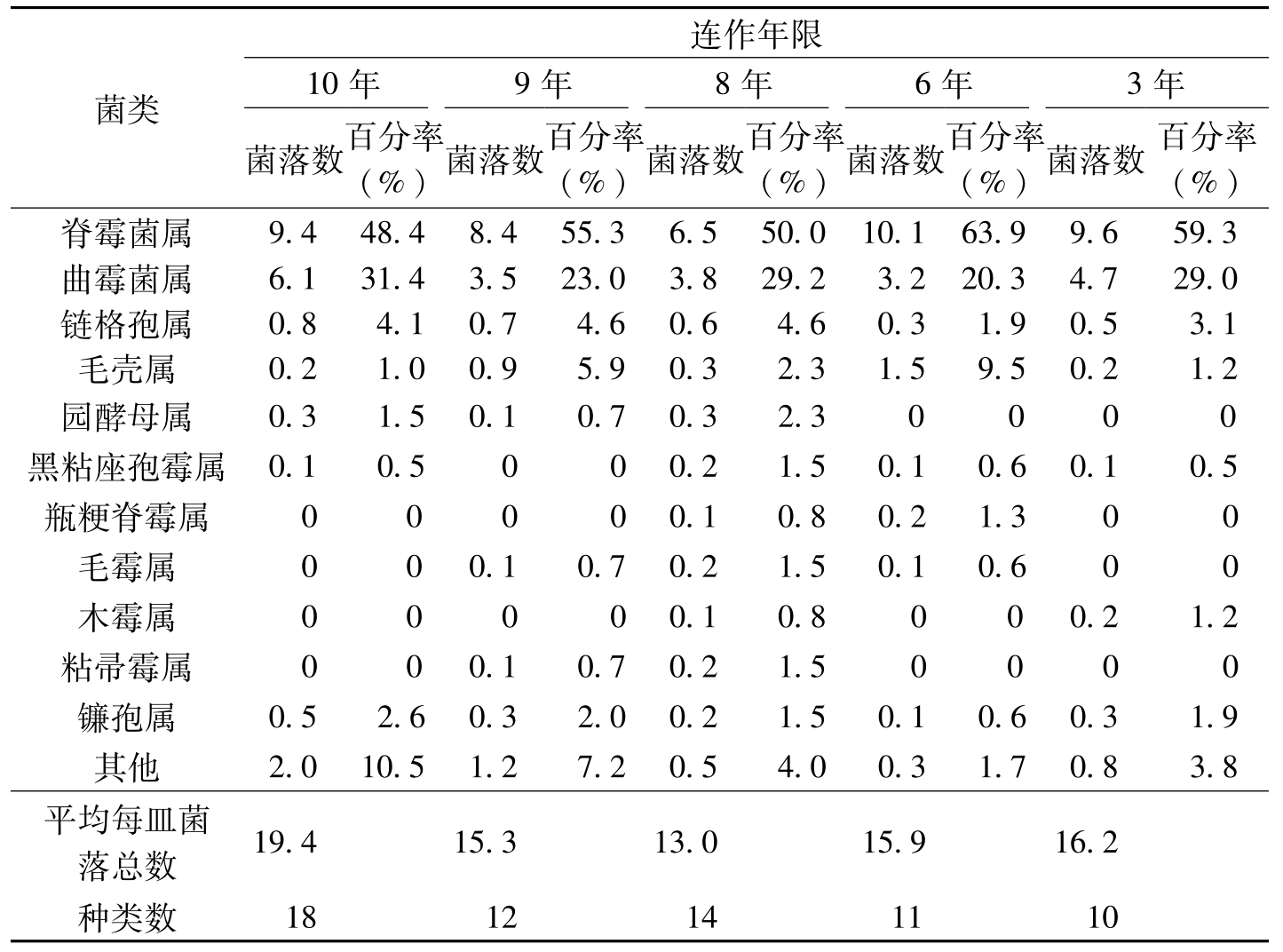
表8-21 棉花不同连作年限土样中真菌类群比较(平均每皿菌落数)
菌类连作年限10年9年8年6年3年菌落数菌落数菌落数菌落数菌落数纤维螺旋链霉菌5456341325产二素链霉菌482323813天冬素紫色链霉菌3725281110球孢链霉菌黄色变种2118141110土味链霉菌48173紫色链霉菌01000球孢链霉菌01000
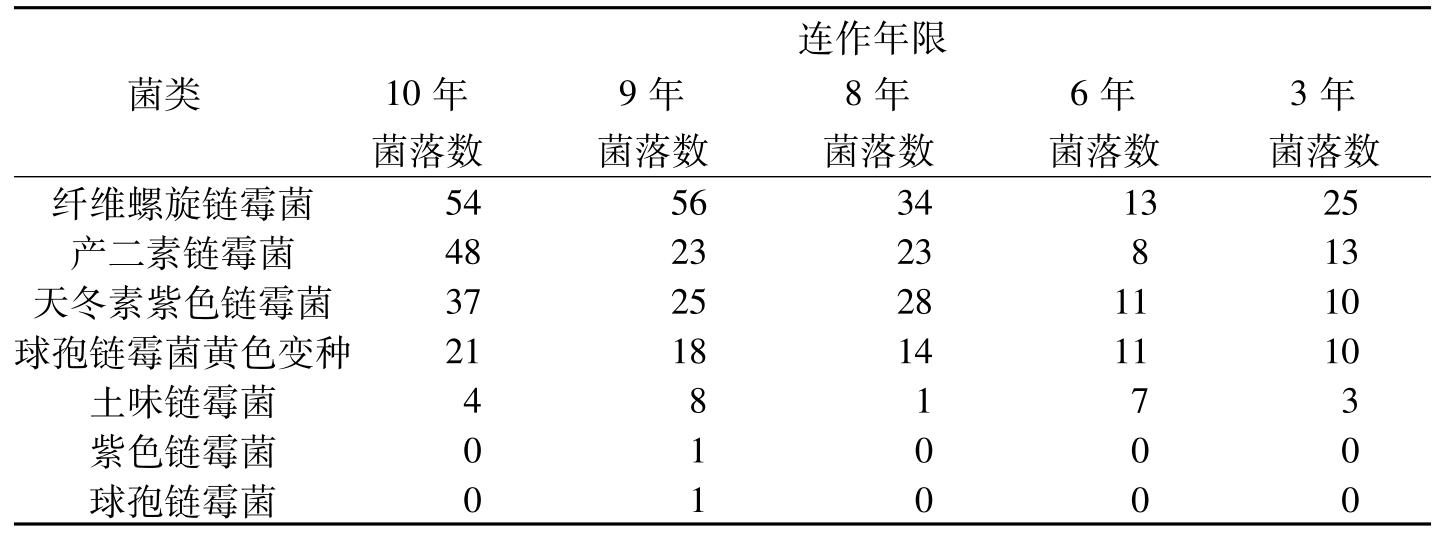
表8-22 不同连作年限土样中放线菌数量的变化(个)
菌类连作年限10年9年8年6年3年菌落数菌落数菌落数菌落数菌落数细黄链霉菌00300绿色产孢链霉菌00200其他6127116合计1701391126167群落数11101197菌落数(皿)28.323.218.710.211.2抑制性菌落数3.53.33.21.81.7
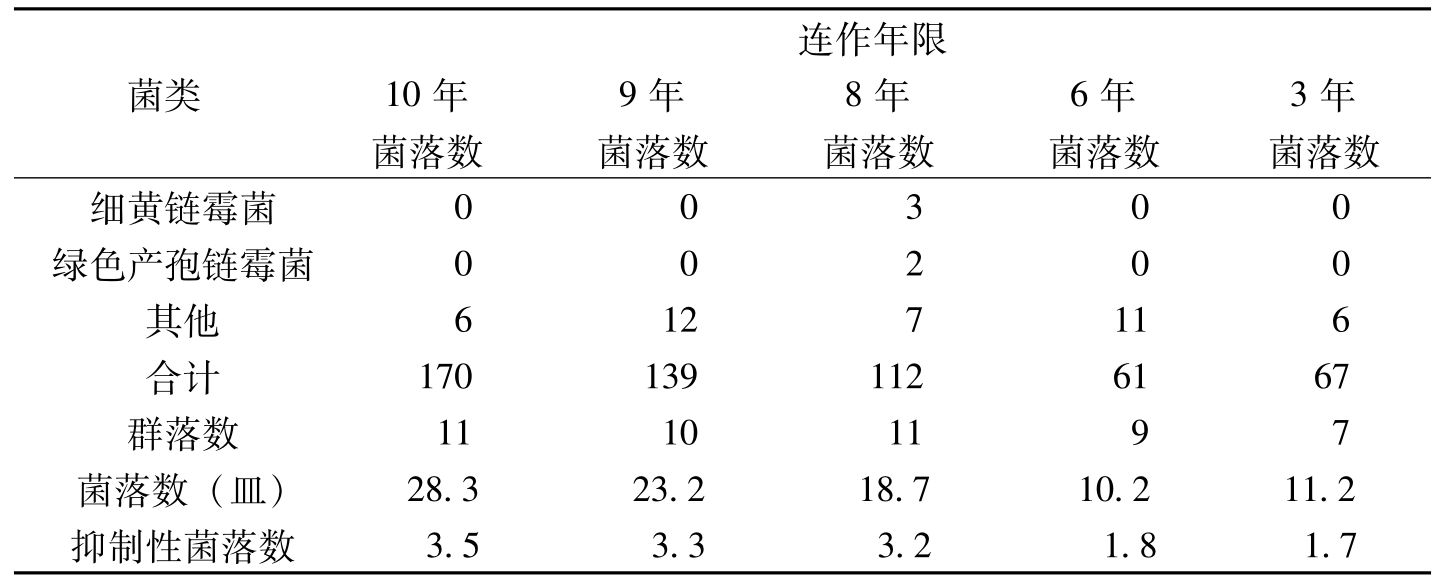
表8-22 不同连作年限土样中放线菌数量的变化(个)(续)-1
菌类连作年限10年9年8年6年3年菌落数菌落数菌落数菌落数菌落数Ⅰ2254596151Ⅱ2925465781Ⅲ25012109Ⅳ00013Ⅴ004130Ⅵ16213Ⅶ011000Ⅷ024000Ⅸ002800菌落数45665菌落数(皿)77120151142153抑制性菌落数25401089
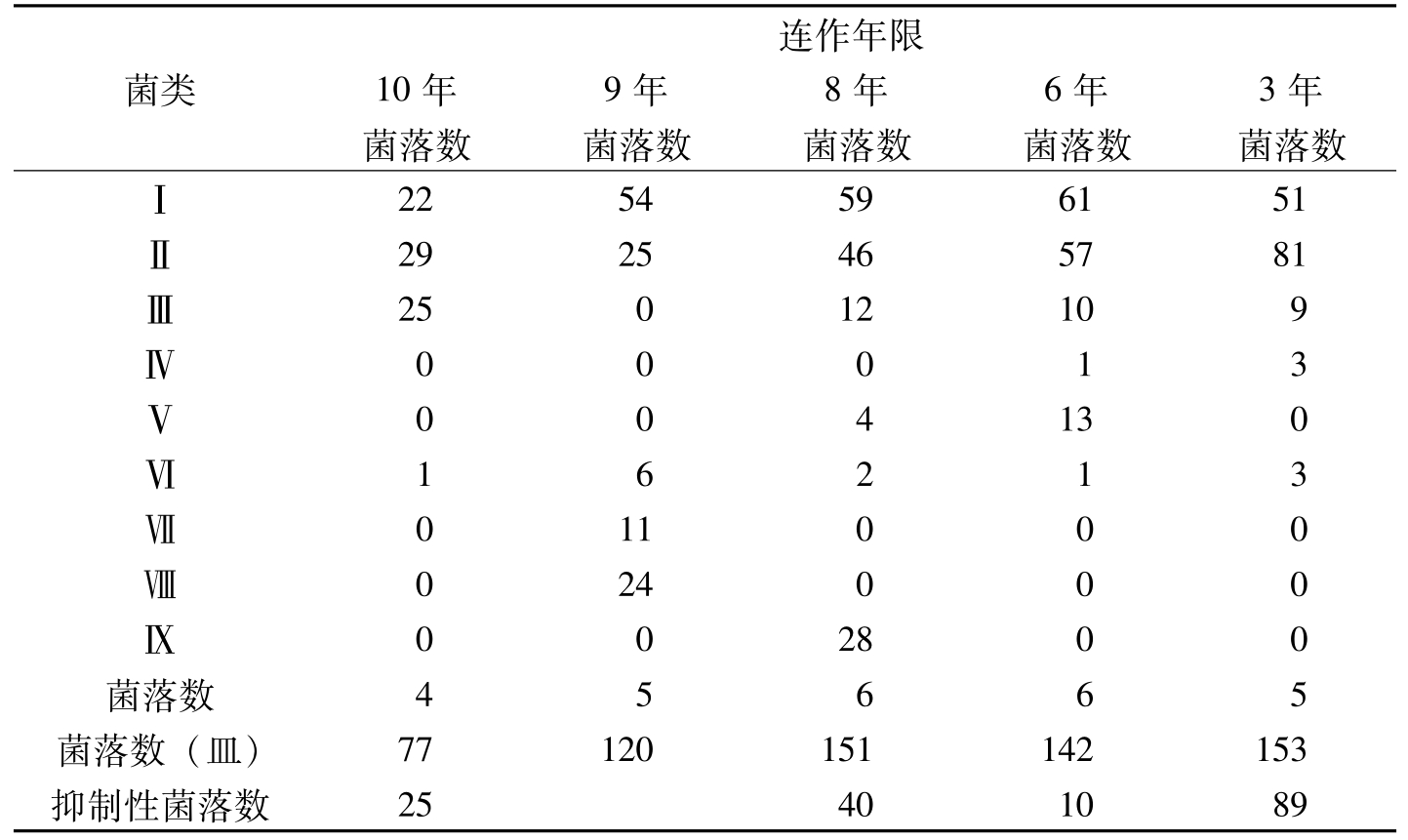
表8-23 不同连作年限土样中细菌的类群及数量(个)
土样螺旋线虫滑刃线虫真滑刃线虫垫刃线虫短体线虫其他合计连作9年16271230058连作6年2014851250连作4年1410028连作2年0110002

表8-24 不同连作年限土样中线虫数量的变化(个)
李长兴等(1996)连续3年对4类型土壤采土,在不同棉花生育期采
土样类型真菌(×103个/克土)细菌(×104个/克土)青霉曲霉木霉有芽孢无芽孢放线菌(×104个/克土)抑菌土(10年)105.869.70.768.3285.8255.8抑菌土(5年)100.360.5047.7204.9221.1小麦田土63.321.2046.3149.6109.9未植过棉土67.830.8041.298.2141.3
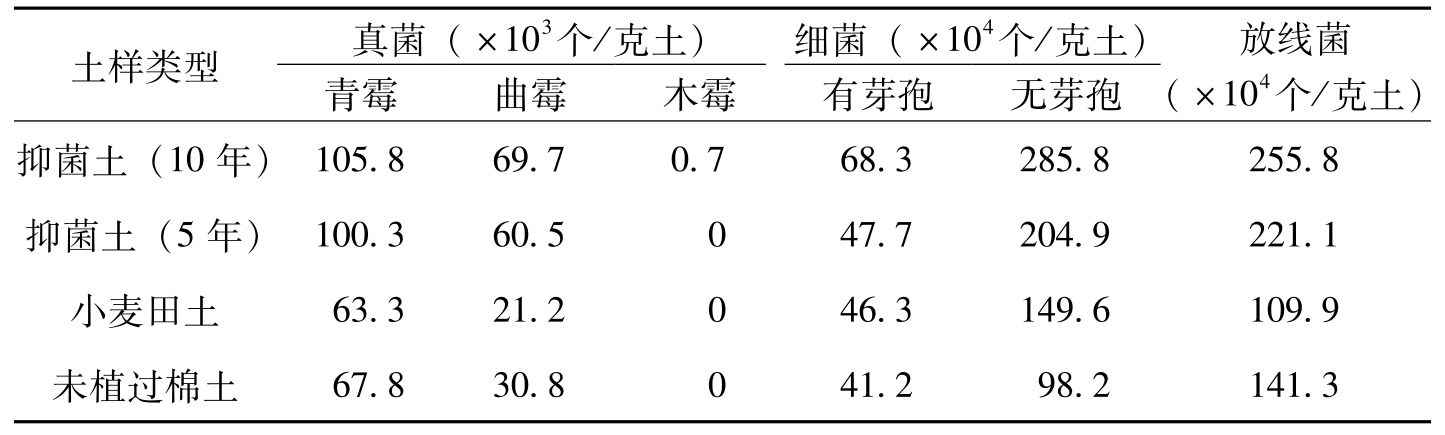
表8-25 不同类型土样菌量
3.抗病品种连作棉田土壤浸出液对病菌的影响
对病菌小孢子萌发、厚垣孢子产生数量的测定结果表明(表8-26),不同连作年限土样浸出液对病菌小孢子萌发的影响没有显著差异;对厚垣孢子的形成却有一定影响:连作6年以下每视野(16×10)平均17~18.8个厚垣孢子,9年有13.3个厚垣孢子,连作15年仅有4.9个厚垣孢子,说明随抗病品种种植年限的增长,厚垣孢子的形成数量逐渐减少,由此推测土壤中,有抑制厚垣孢子形成的物质,该物质随年限的增长而逐渐增多。
土样来源病菌小孢子萌发率(%)病菌厚垣孢子的形成数量(个/视野)ⅠⅡⅢⅣ显著水平*松林土(CK)81.120.224.311.818.8a重病田土70.720.514.217.817.5a连作6年70.720.718.517.318.8a连作9年63.915.017.87.213.3ab连作11年—11.27.012.810.3ab连作15年84.63.37.83.74.9b
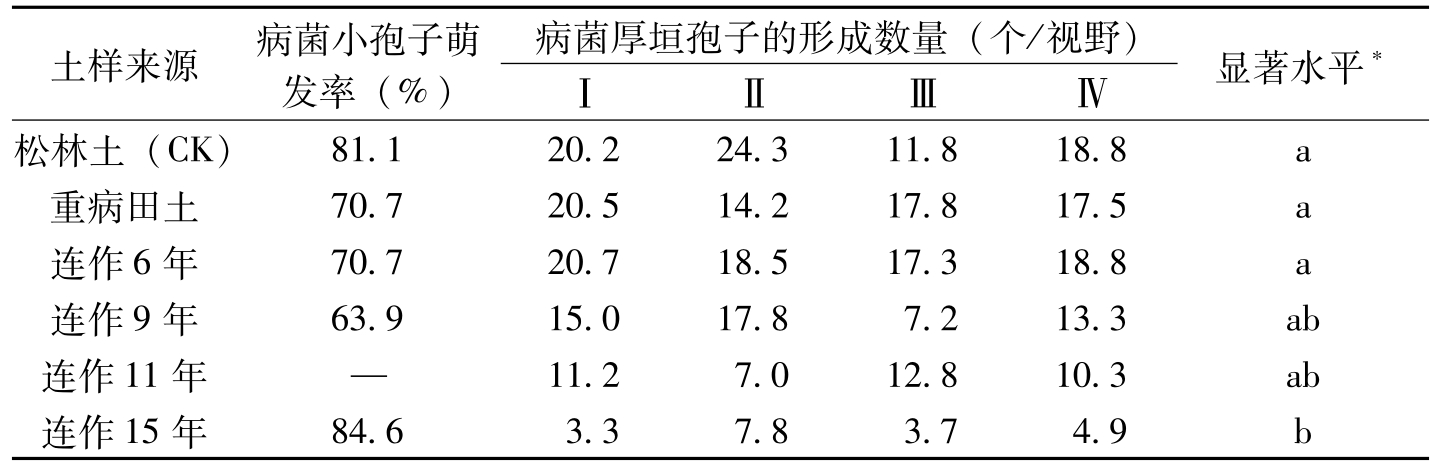
表8-26 不同土样浸出液对棉枯萎菌的影响(杨之为等,1995)
总之,对棉花枯萎病抑菌土的研究,为枯萎病等多种病害开辟了生物防治途径,利用抑菌土或筛选抑菌生物因子防治植物病害,随着研究的不断深入和技术方法的逐步完善,将可能取得较大的经济效益和社会效益。
尽管棉花枯萎病的生物防治研究工作已历时数十年,但至今未形成商品化制剂。究其原因,马存等(2007)认为,是由于对许多问题还缺乏研究,许多困难有待解决。与整个生物防治普遍存在的问题一样,在这些问题与困难解决之前,生物防治尚难以作为常规手段用于棉花枯萎病的治理。这些问题主要包括:一是生态学障碍。从实验室向大田阶段的过渡往往遭到失败,其中,生态障碍是主要原因;二是防治对象单一。生物防治研究仅仅针对其中一种或几种病害,往往不被工业化生产所接受;三是缺乏适合工业化生产的扩繁技术。在实验室内可以不计成本培养所需生防制剂,但在商业化生产上则要考虑经济效益,与规模化生产有关的大量培养技术则有待提高;四是缺乏准确可靠的筛选技术与筛选标准。目前,生防菌的筛选技术和标准仍然是沿用传统的平板对峙法和在平板上对病原菌的拮抗能力,其预见性较差,如果结合生防菌的其他特性,如定殖能力的测定,则有望筛选到更有潜力的菌株;五是缺乏规范化的菌剂制备技术。有关生防菌剂制剂化和规范化的配套研究相对缺乏,无法将有效的生防菌开发成规范的产品;六是缺乏配套的生防菌田间施用技术。植病生防的研究目的是防治田间植物病害,其田间施用技术选择的适当与否,决定着该菌剂的有效性。针对上述棉花枯萎菌生防菌所存在的问题,结合工业化生产需要,认为生防菌株应具有如下特点:①生长速度快,产孢量大。②作用谱广泛。③作用机制多样性。④可在棉花根际大量定殖。⑤能作用于棉花内部的枯萎病菌。⑥不受土壤抑菌作用的干扰。⑦休眠孢子有较强的耐干燥能力。⑧对棉花生长有促进作用。⑨要有合适的生防菌剂剂型及相应的施用方法。虽然目前没有登记注册的防治棉花枯萎菌的生防菌,但已有防治其他作物枯萎病的生防制剂商品。例如,意大利S.I.A.P.A公司登记的利用非致病性尖孢镰刀菌作为生防菌的产品Biofox C,可防治由尖孢镰刀









