Research Advances on the Enterotoxin of the Bacillus cereus
芽孢杆菌属(Bacillus spp.)是一类好氧或兼性厌氧、产生抗逆性内生孢子的杆状细菌,许多为腐生菌,主要分布于土壤、植物体表面及水体中,其在工业、农业、医学、军事和科学研究中有广泛的应用价值。
在《Bergey氏鉴定细菌学手册》第8、第9版中,蜡样芽孢杆菌的分类地位为芽孢杆菌属的第I群,该群有22个种。根据营养型菌细胞的宽度分为两类,蜡样芽孢杆菌、蕈状芽孢杆菌、苏云金芽孢杆菌、炭疽芽孢杆菌和巨大芽孢杆菌属“大细孢菌种”。
1 蜡样芽孢杆菌的危害? 项目来源:国家自然科学基金(30640044)。
蜡样芽胞杆菌是一种杆状、产内生芽孢的革兰氏阳性细菌,由于蜡样芽孢杆菌自然界分布甚广,常存在于土壤、灰尘、腐草和空气中,极易在食品加工、运输、贮存、销售过程中,通过苍蝇、蟑螂等昆虫和不卫生的用具和手污染,通常被认为是一种条件致病菌,在临床上可导致脓肿、脑膜炎、骨髓炎、心内膜炎等报道,但最常见的是导致两种不同类型的食物中毒:腹泻型和呕吐型。
关于B.c.引起非肠道感染及食物中毒的例子很多,从1898年起,就有B.c.造成泌尿系统感染及肠胃炎的记载,有些感染的病例甚至很严重,以致造成死亡。在微生物发展的早期,好氧芽孢杆菌就被怀疑可造成食物中毒,Lubenau1906年描述了发生在一家医院的严重的食物中毒事件,300名医务人员及病人用餐后出现急性肠胃炎,对剩余的食物进行检测,发现含有大量的好氧芽孢杆菌,该污染菌为B.c.。Seitz 1913 年从一例患肠炎与腹泻的病人分离出B.c.。Brekenfeld 分别于1926年及1959年报道了两起B.c.造成的食物中毒事件。1936~1942年,瑞典卫生部对367例食物中毒事件综合分析,证实117例是由B.c.引起,并且认识到被B.c.污染的食物,储藏温度不当时,可能会造成食物中毒,在1973年Bulyba等人报道了污染蜡样芽孢杆菌的乳制品引起食物中毒。由于Smith、Gorden 及其同事在芽孢杆菌分类学上的进展,Hauge 经过对4起食物中毒事件的调查,于1995年首次确认B.c.是一种引起食物中毒的致病菌。
目前大部分国家对各类食品中的蜡样芽孢杆菌数量有所限定,多数情况下,引起食物中毒的食品中蜡样芽孢杆菌的数量在105~108 CFU/g,常因食用肉类、海鲜、乳品和蔬菜等食物引起,潜伏期一般为6~15h,一般持续24h;而致呕吐的毒素是该菌在食物中预先产生的,该毒素非常稳定,进入人体后在胃中与其受体5-HT3 结合,导致呕吐。呕吐型食物中毒的潜伏期一般为0.5~6h,一般限于富含淀粉质的食品,特别是炒饭和米饭。主要症状为恶心、呕吐,有时有腹泻、头晕、发烧和四肢无力等症状,引起这两种食物中毒的食品通常都是经过热加工处理的,但蜡样芽孢杆菌具有耐热的芽孢,能在食品加工及烹饪后残留下来,热处理诱发芽孢的萌发,在没有其他微生物与之竞争的条件下,大量生长繁殖,产生毒素并引起食品的腐败。
2 蜡样芽孢杆菌产生的毒素
2. 1 呕吐型毒素
蜡样芽孢杆菌产生的呕吐毒素(cereulide,1.2kD)是一种小的十二边形的热稳定性环状毒素,分子式为(D-O-Leu-D-Ala-L-O-Val-L-Val)3。其结构、性质和毒理与缬氨霉素很相似,是特异性的钾离子载体,能将K 转入线粒体内,破坏线粒体的氧化还原能力。该毒素非常稳定,目前的各种食品加工方法,包括灭菌,均无法使其失活(能耐受126℃ 90min),而且还耐强酸(pH 2.0)、耐蛋白酶水解。N.Agata等对多种食品中呕吐毒素的产量进行了检测,发现对B.cereus NC7401来说,在煮熟后的米饭中其产毒量很高,在富含淀粉质的食物中的产毒量也足以引起食物中毒;而在肉类、蛋品和密封的液体食品如牛奶和豆奶中虽可以检测到该毒素,但其含量较低。还发现在与醋、蛋黄酱及酱类一起煮的食物中,该菌株的生长和产毒都受到抑制,推测这可能是醋导致pH 降低的缘故。在12~15℃时该毒素的产量却明显高于30℃时的产量,而且该毒素的产生与芽孢的产生没有相关性。还有报道称该毒素只有在有氧条件下才能产生,所以缺氧条件如:充氮包装和真空包装能有效地防止该毒素的产生和积累。因为该毒素的分子量很小,无抗原性,这使其检测比较难,到目前为止尚缺乏一种快速可靠的检测方法。最常用是采用HEp-2 细胞进行细胞培养分析。近年来用分子生物学手段检测产毒菌株的报道也较多,如P F Horwood 等人根据NRPS基因的两个可变区的序列,针对产呕吐毒素的菌株设计了特异性的引物,进行PCR 以检测蜡样芽孢杆菌是否产毒,取得了良好的效果,该法灵敏度高,而且检测速度快。在呕吐食物中毒事件中分离的蜡样芽孢杆菌均产生呕吐毒素,而且有着共同的独特表型特征,对其基因进行分析发现它们同源性很高。
2. 2 腹泻型毒素
B.c.所造成的腹泻型食物中毒的致病因子是肠毒素,目前至少已经发现4 种不同的肠毒素,包括2 个三联体肠毒素:溶血素BL和非溶血素Nhe;2个单一亚基肠毒素:细胞毒素K(cytK)、肠毒素T(bceT)。
2.2.1 溶血素BL(HBL)Beecher1991年从B.c.菌株中分离提纯了一种具有活性的三亚基肠毒素,命名为溶血素BL(hemolysinBL),能够引起家兔肠段的液体积累,可以改变豚鼠皮肤血管的通透性,具有对vero细胞的溶细胞毒性。其由一个结合亚基B(37.5kD)、两个溶血亚基L1(38.2kD)及L2(43.5kD)组成,编码3个亚基的基因hblA、hblD、hblC经克隆、测序与分析,表明其在同一个mRNA中受一个操众子调控转录如图1。这些组分的物理化学性质非常相似,等电点(pI)为5.34,5.33和5.33。其中hblA编码结合亚基B,hblD、hblC分别编码溶血亚基L1及L2,hblB编码B’蛋白,hblC和hblD仅隔37bp个碱基,B、L1、L2蛋白分别有31、30、32个氨基酸的信号肽,hblD和hblA之间最少有100bp碱基,hblA和 hblB有381 bp的碱基隔开,B’蛋白与B蛋白开始的158个氨基酸非常相似,但其功能尚未清楚。Douglas J.Beecher等人利用等电聚焦电泳技术和快速蛋白液相层析技术证明单独成分的溶血素亚基并不会在血平板上产生溶血环,只有当3个亚基结合后,才会产生溶血环。
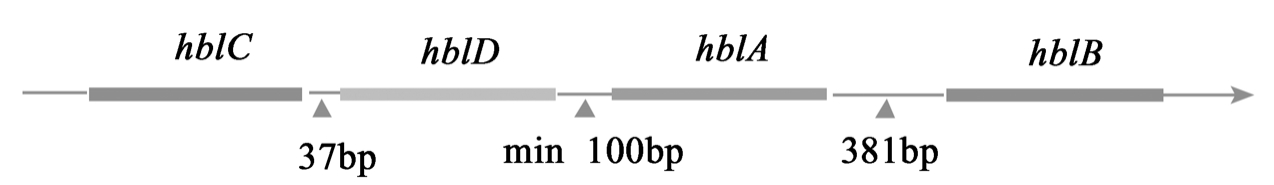
图1 芽孢杆菌溶血素BL操纵子图谱
2.2.2 非溶血素肠毒素(Nhe)非溶血素肠毒素(Nhe)由45、39和105kD的蛋白组成,其蛋白成分已被分离出来。1999年Granum等给出了nhe操纵子的序列,该操纵子有3个开放式阅读框,相应的3个基因分别是:nheA、nheB和nheC。前两个基因的产物分别为45kD和39kD 蛋白,而nheC 的产物尚未纯化出来,其功能未知。Nhe与Vero细胞相互作用的研究表明105kD蛋白是复合物的结合部位,而其他两个组分是无法单独结合到细胞上去。该105kD 蛋白是一种金属蛋白,具有分解明胶和胶原的活力。与hbl 基因不同,编码该毒素的基因位于质粒上。
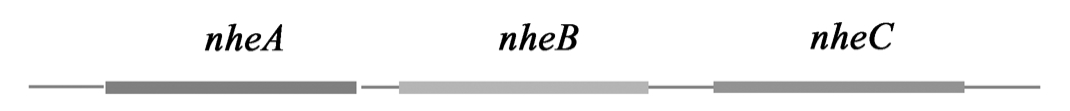
图2 芽孢杆菌非溶血素Nhe操纵子图谱
溶血素BL(HBL)与非溶血素肠毒素(Nhe)同时受到PlcR的调控。
2.2.3 肠毒素T(entertoxin T)肠毒素T为单一亚基的蛋白质,由 bceT基因编码,日本学者Agata对其基因克隆、测序和分析表明其由336个氨基酸组成。并认为其有细胞毒性,可导致家兔肠段的液体积累,可以改变豚鼠皮肤血管的通透性,具有对vero细胞的溶细胞毒性。其产物属于肠毒素蛋白。该毒素同溶血素BL无同源性,而且认为肠毒素T 不会导致食物中毒。
2.2.4 细胞毒素K 早期在法国报道过食物中毒,其氨基酸序列显示它属于β-桶孔形成毒素,能在磷脂双分子层中形成直径至少为7A°的孔,该孔具有微弱的离子选择性,已证实它对人类肠道Caco-2上皮细胞具有毒性。
3 蜡样芽孢杆菌毒素的调控
PlcR是条件性人类病原菌B.cereus和共生病原菌B.thuringiensis细胞外毒性因子的一个多效调节子,它在细胞进入稳定期时诱导生长。受到PlcR调节的基因有:plcA编码一个专一性磷脂酰肌醇磷脂酶C(PI-PLC),Plc编码一个改良的磷脂酰胆碱磷脂酶 C(PC-PLC),nhe编码一个无溶血性的肠毒素,hbl编码一个溶血性的肠毒素 BL(HBL);以及推定为S-层类似表面蛋白的基因,以及一个推定为细胞外RNA酶。通过分析37.1kb的hbl,plcA和plcR周围的DNA序列,推定存在28个ORF。3条新基因推定受到PlcR 的调节并编码一个中性蛋白酶,subtilase家族丝氨酸蛋白酶(Sfp)以及一个推定的细胞壁水解酶(Cwh)得到确认。相应的sfp和cwh 基因定位于plcA的上游调节区域,能同时受到位于逆转录基因之间的PlcR结合位点的调控。
Sylvie Salamitou等构建plcR基因缺失的突变菌株,该基因编码一个多效细胞外因子的调节子。幼虫期同时取食亲本菌株产生的106孢子亚致死浓度的Cry1C毒性导致70%死亡率,如果使用plcR突变体的孢子,则只有7%的死亡率。小鼠鼻腔灌入108的孢子,亲本菌株导致了100%的死亡率,而灌入相同数量的突变体孢子,死亡率大大降低,甚至没有死亡。应用营养体细胞代替孢子也可以达到相同的效果。导致死亡的原因未知,不可能是由于小鼠内细菌的实际增长所导致。由于受B.thuringiensis 过量突变体感染的小鼠产生的病变,说明溶血素参与其中,发生了作用。B.thuringiensis和B.cereus具细胞溶解毒性的特性。这种细胞溶解毒性的水平在plcR基因缺失的菌株中剧烈下降。表明 B.thuringiensis407菌株和B.cereusATCC14579的致病性受到PlcR的调控。
4 蜡样芽孢杆菌毒素的检测
由于蜡样芽孢杆菌及其芽孢广泛存在于周围的环境中,它极易污染食物而引起食物中毒,因此需要发展一种快速的检测方法来实现对致病性蜡样芽孢杆菌的检测,目前对该类蜡样芽孢杆菌的检测主要采用生化检测方法是一项费时费力的方法,需要长时间的选择性培养过程。现在有两种试剂盒可供选择,但由于价格昂贵且不太灵敏,有些致病菌不能够检测。王利国等人对实验室14株芽孢杆菌溶血素BL的检测结果表明,8株蜡样芽孢杆菌全部检测到溶血素BL的基因且产生溶血环,而其他的蜡样芽孢杆菌只检测到hblA基因以外的基因且不产生溶血环,表明只要检测到hblA基因,证明其为致病性菌株,所以通过设计hblA基因特异引物用PCR或通过血平板培养的方法是既经济又快速的检测方法。对其他毒素的检测目前主要是通过设计特异性引物来检测。所以对蜡样芽孢杆菌毒素的检测还需要进一步对其研究,确定最佳的检测方法。









