Nitric Oxide and Hydrogen Peroxide Signaling in Tobacco Resistance to Tobacco Mosaic Virus Induced by Oligogalacturonic Acid
1 Introduction
Plant and fungal cells are surrounded by a cell wall rich in diverse polysaccharides and proteins.It has become apparent in recent years that the carbohydrates in the cell wall function not only to maintain cell shape and integrity,but also may serve as signals in plants(Mohnen et al.,1993).
Oligogalacturonic acid(OGA),a well studied elicitor,is derived from plant cell walls(Nothnagel et al.,1983).When added to cultured plant cells,it induces an oxidative burst within minutes,releasing ROS via a pathway that involves receptor binding,activation of a G-protein,influx of Ca2+,stimulation of phospholipase C,and induction of a number of kinases(Apostol et al.,1989;Horn et al.,1989;Legendre et al.,1992;Chandra et al.,1995;Legendre et al.,1993).Purified OGAs 13 to at least 26 residues long stimulate pp34 thiophosphorylation in vitro(Philippe et al.,1995).OGAs are also involved in the induction of the jasmonate pathway during plant defense response to E.carotovora subsp.Carotovora attack(Cecilia et al.,1999).
The first response observed after the addition of OGAs that is clearly involved in plant defense is the production of active oxygen species,including H2O2,and O2-.This response,termed the oxidative burst,occurs within a few minutes after the addition of OGAs to suspension-cultured soybean,tobacco,and tomato cells.Reactive oxygen species are thought to have direct(through cytotoxicity)and indirect(through signaling)roles in the plant cell death required for the HR.Reactive oxygen species induce the expression of defense related genes,and are implicated as second messengers that elicit other defense responses,including systemic acquired resistance(SAR)and the HR(Brent etal.,2001).
Different elicitors are thought to activate different sets of second messengers.The two signaling events that appear to participate in the OGAs inducing plant defense include the oxidative burst and NO accumulation.Inhibitors of mammalian nitric oxide synthase reduced both OGA-induced NO ac-cumulation and NOS activity,suggesting that OGA-induced NO production occurs via a NOS-like enzyme(Hu et al.,2003). Nitric oxide(NO)is a highly reactive molecule that rapidly diffuses and permeates cell membranes.During the last few years NO has a significant role in plant resist-ance to pathogens by triggering resistance-associated cell death and by contributing to the local and systemic induction of defense genes.NO stimulates signal transduction pathways through protein ki-nases,cytosolic Ca2+mobilization and protein modification(María et al.,2004). Most of the ex-perimental data available on NO detection during plant-pathogen interactions come from studies of infections by biotrophic pathogens(María et al.,2004). Additionally,an increase in NOS activity correlated with the pathogen resistance response has been observed in resistant tobacco during TMV infection( Durner et al.,1998;Chandok et al.,2003).
Here we report that OGAs induced a range of defense responses in tobacco,including oxidative burst,NO accumulation and stimulation of superoxide dismutase(SOD)activity and catalase(CAT)activity.Furthermore,we show that tobacco plant sprayed with OGAs developed a resistance against infection by tobacco mosaic virus.We also provide evidence that the defense response induced by OGAs was connected with H2O2 and NO pathway.
2 Materials and methods
2.1 Plant materials
Plants of tobacco(Nicotiana tabacum var.sam sun NN)were grown from seeds in a greenhouse and were used at the 4~6-leaf stage after 2 months in culture.The plants were kept in a growth chamber at(23±1)℃ with a photoperiod of 16 h and 70%~80%relative humidity for several days before treatments.
2.2 Chemicals
Diphenylene iodonium(DPI),2-(N-morpholino)ethanesulfonic acid(MES),Sodium nitroprusside(SNP),catalase(CAT,from bovine liver),NG-nitro-L-arginine-methyl eater(L-NAME)and 4,5-diaminofluorescein diacetate(DAF-2 DA)were obtained from Sigma.2′,7′-dichlorofluorescin diacetate(H2DCF-DA)from Biotium.All other reagents were from Shanghai Chemical Reagent CO.,LTD,Tianjin Kermel Chemical Development Centre,or Beijing Chemical Plant.
2.3 Preparation of OGAs
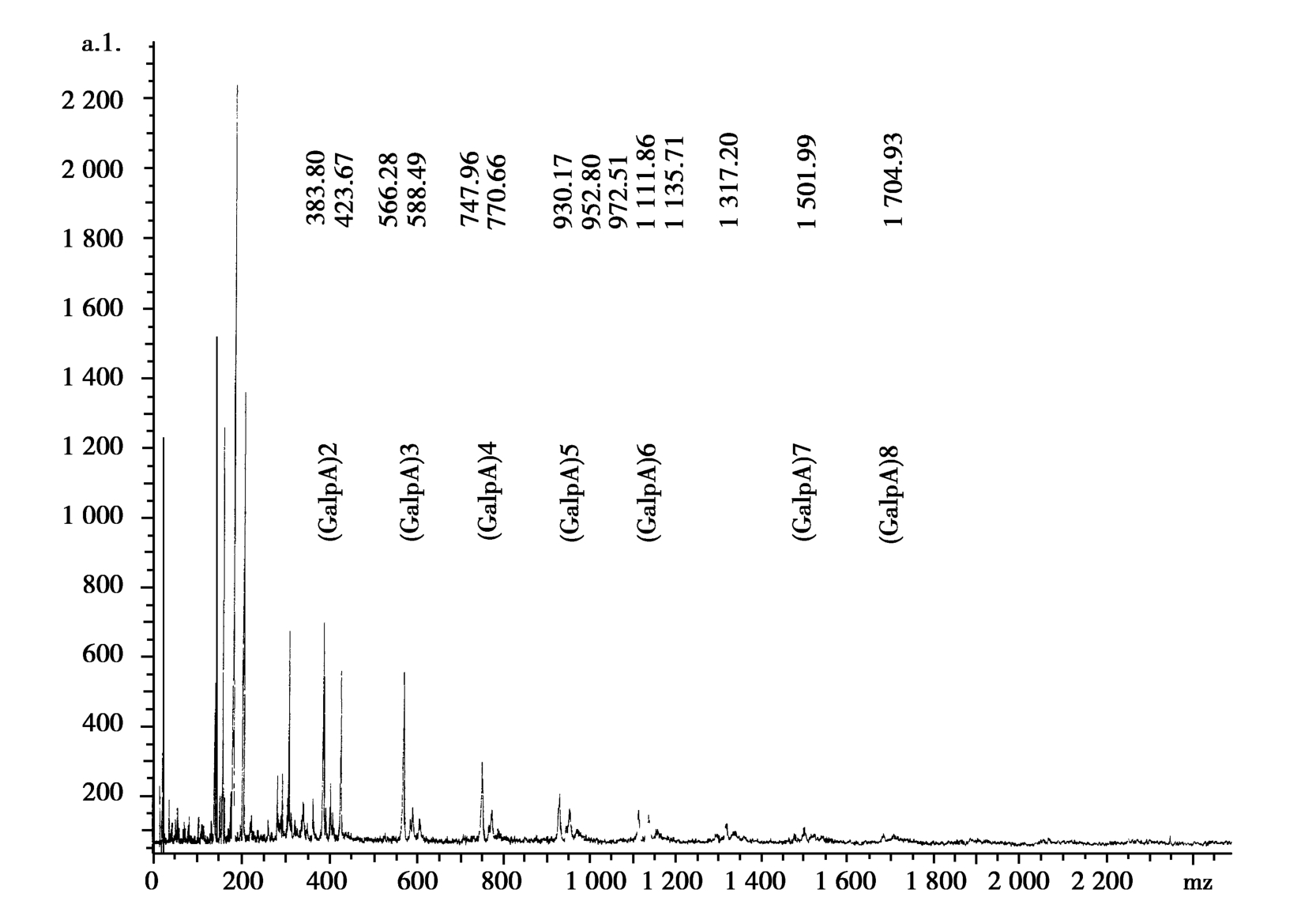
OGAs was prepared from enzymatic hydrolysis of pectin and separated with membrane according to the report(H Zhang et al.,1999).An aliquot of OGAs was dissolved in water and analyzed with a matrix-assisted laser desorption-ionization time-of-flight mass spectrometer(MALID-TOF-MS,Bruker,Germany).
2.4 Assay for effect of OGAs on controlling TMV
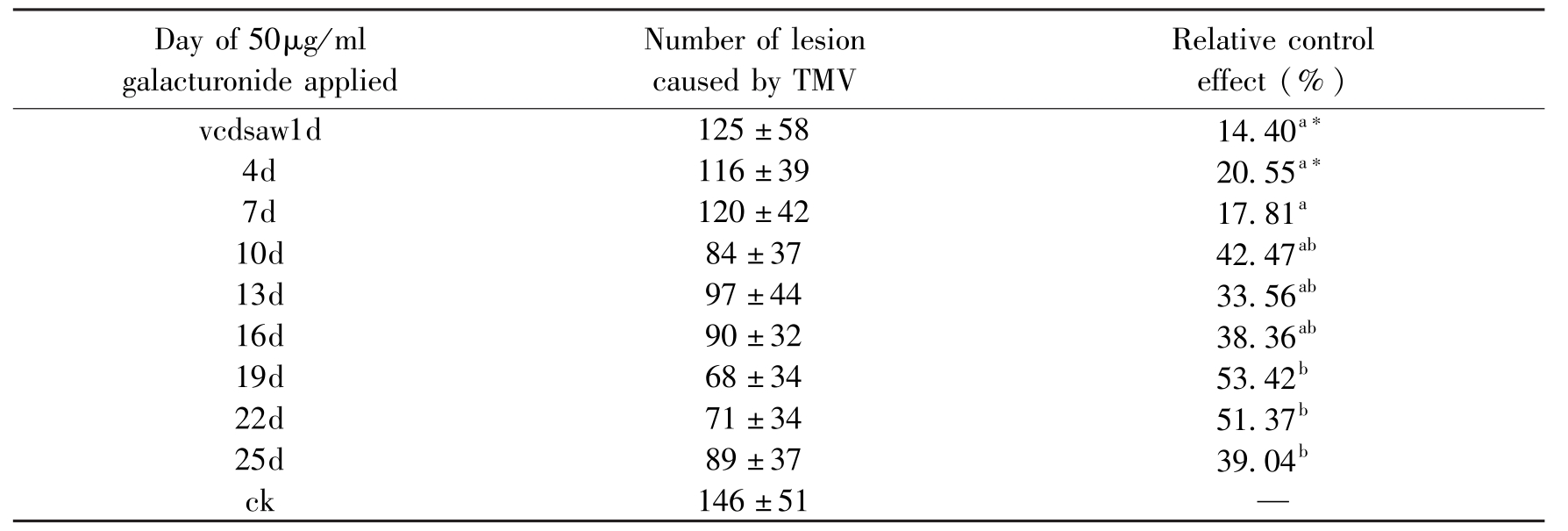
Tobacco mosaic virus(TMV)that came from our collection was multiplied in N.tabacum.TMV was extracted from systemic infected plants by homogenization of infected leaves in 0.05mol/LH3PO4 buffer(0.05mol/L KH2PO4,0.05 M Na2HPO4 pH 6.8)with subsequent clarification of the extract by centrifugation at 2000g for 6 min.The supernatant extract was used for mechanical inoculation.All leaves of plant were sprayed with 50μg/ml of OGAs,the control plants were sprayed with water.24h~25d after OGAs application,plants were inoculated mechanically with TMV.The lesion caused by TMV was investigated at 7d after inoculation.Results were analyzed using Duncan's multiple range test at P= 0.05.
2.5 Enzyme extraction and superoxide dismutase (SOD) and catalase (CAT) assay
For measurements of SOD and CAT activities,tobacco leaves treated with OGAs were kept in liquid nitrogen.The enzymes in the frozen powders were extracted by adding 0.05g polyvinylpyrrolidone and 5ml 0.05mol/L sodium borate buffer at pH 8.8 and homogenized at 4℃.SOD activities were measured as described by Zhu Guanglian(Zhu Guanglian et al.,1990).CAT activity was determined using the method of Beers&Sizers(Beer et al.,1952).
2.6 Measurement of production of NO and H2O2 in epidermal tobacco cells by laser scanning confocal microscopy
NO and H2O2 measurement was performed using their fluorescent indicator dye DAF-2 DA and H2DCF-DA as described previously by H.Kojima(H.Kojima et al.,1998)with slight modifications.The epidermis was peeled carefully from abaxial surface of the leaves and cut into 5-mm length.Epidermal strips were placed into Tris/KCl buffer(Tris 10 mmol/L and KCl 50mmol/L,pH 7.2)containing DAF-2 DA at a final concentration of 10μmol/L for 30min,or H2DCF-DA at 50μmol/L for 10min,at 26℃ in the dark.The epidermal sections were removed and transferred to a dish of fresh Tris/KCl buffer(without probe)to wash off excess fluorophore apart from light.Then the epidermal strips were placed in Tris/KCl buffer containing OGAs and inhibitors.Examination of peels was performed using laser scanning confocal microscopy(Leica,TCS SP2)with exciting wavelength 488 nm,emitting wavelength 505~530nm.
2.7 Assay for the effect of OGAs and exogenous NO and H2O2 on disease symptom
Plants were sprayed with 0.01 and 0.1 mmol/L of sodium nitroprusside(SNP),50μg/ml of OGAs,1 mmol/L,10mmol/L and 100 mmol/L H2O2,H2O2 scavenger catalase(CAT,100unit/ml)and OGAs cotreatment,H2O2 scavenger ascorbic acid(0.1mmol/L)and OGAs cotreatment and NOS inhibitor L-NAME(1mmol/L)for 30min before OGAs respectively.The control plants were sprayed with water.In all cases,24h after OGAs and other materials applications,plants were inoculated with TMV.The lesion caused by TMV was investigated at 7d after inoculation.The effect of OGAs,SNP and H2O2 on local infection was calculated from the ratio of the number of local lesion produced on the treated leaves to that on the control leaves treated with water.
3 Results
3.1 Preparation of OGAs
The TOF-MS profiles of OGAs sample were showed in Figure 1.The mass spectrum indicated that peaks corresponding to the mass numbers of( M+ Na)+of trimer to enneamer were detected.So the sample was composed mainly of OGAs having degree of polymerization( DP)2-8.
Figure 1 TOF-MS of oligochitosan sample
3.2 Effect of OGAs on local infection of tobacco with TMV
The results of control effects on TMV with OGAs at different concentration(50~100μg/ml)showed that the best concentration was 50 μg/ml(data not shown).The effects of application of OGAs at different time were summarized in Table 1.It was found that tobacco leaves treated with OGAs were protected against TMV infection.When the inoculation occurred at 19d after spraying 50μg/ml OGAs on tobacco plants,the relative control effect was 53.42%.We concluded that the resistance induced by OGAs became better with the inducing time until 19d.The resistance was reduced after 19d.
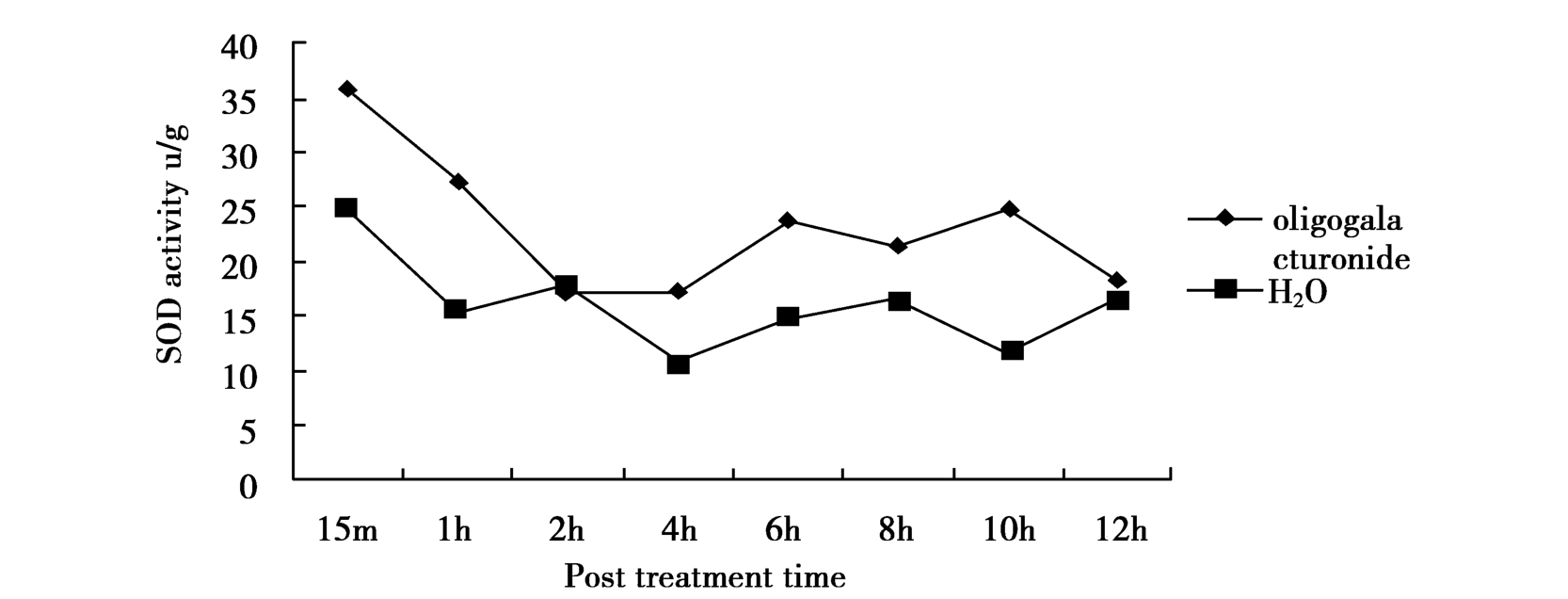
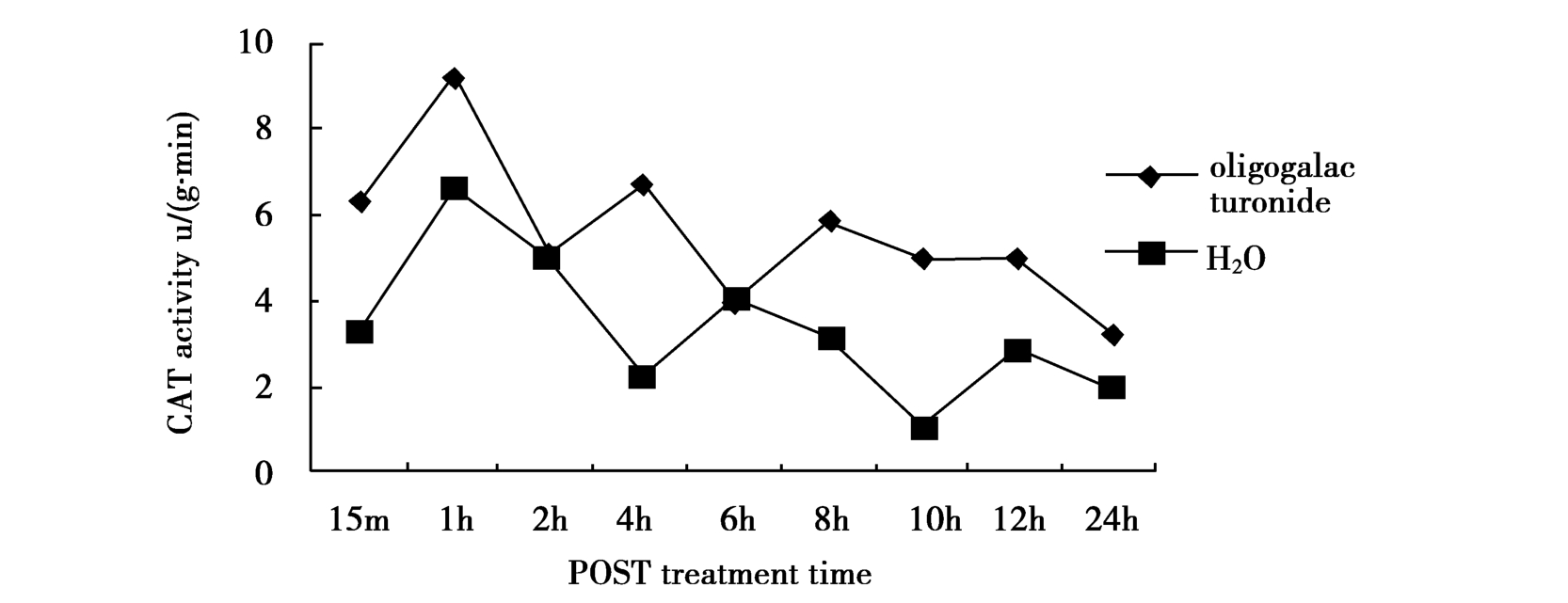
We examined the effects of OGAs on the activity of plant resistance correlated enzymes.The results(Figure 2 and Figure 3.)indicated that OGAs increased activity of SOD and CAT compared with the H2O-treated ones.There are no distinct differences on the activity of POD and PPO of tobacco leaves treated with OGAs or water(data not shown).SOD and CAT are concerned with eliminating oxygen free radical.Within one hour,activities of CAT and SOD were induced to maximum.
Figure 2 Time course of SOD activity in tobacco leaves treated by 50μg/ml OGAs or H2O as CK
Figure 3 Time course of CAT activity in tobacco leaves treated by 50μg/ml OGAs or H2O as CK
3.4 Measurement of production of NO and H2O2 in epidermal tobacco cells by laser confocal scanning microscopy
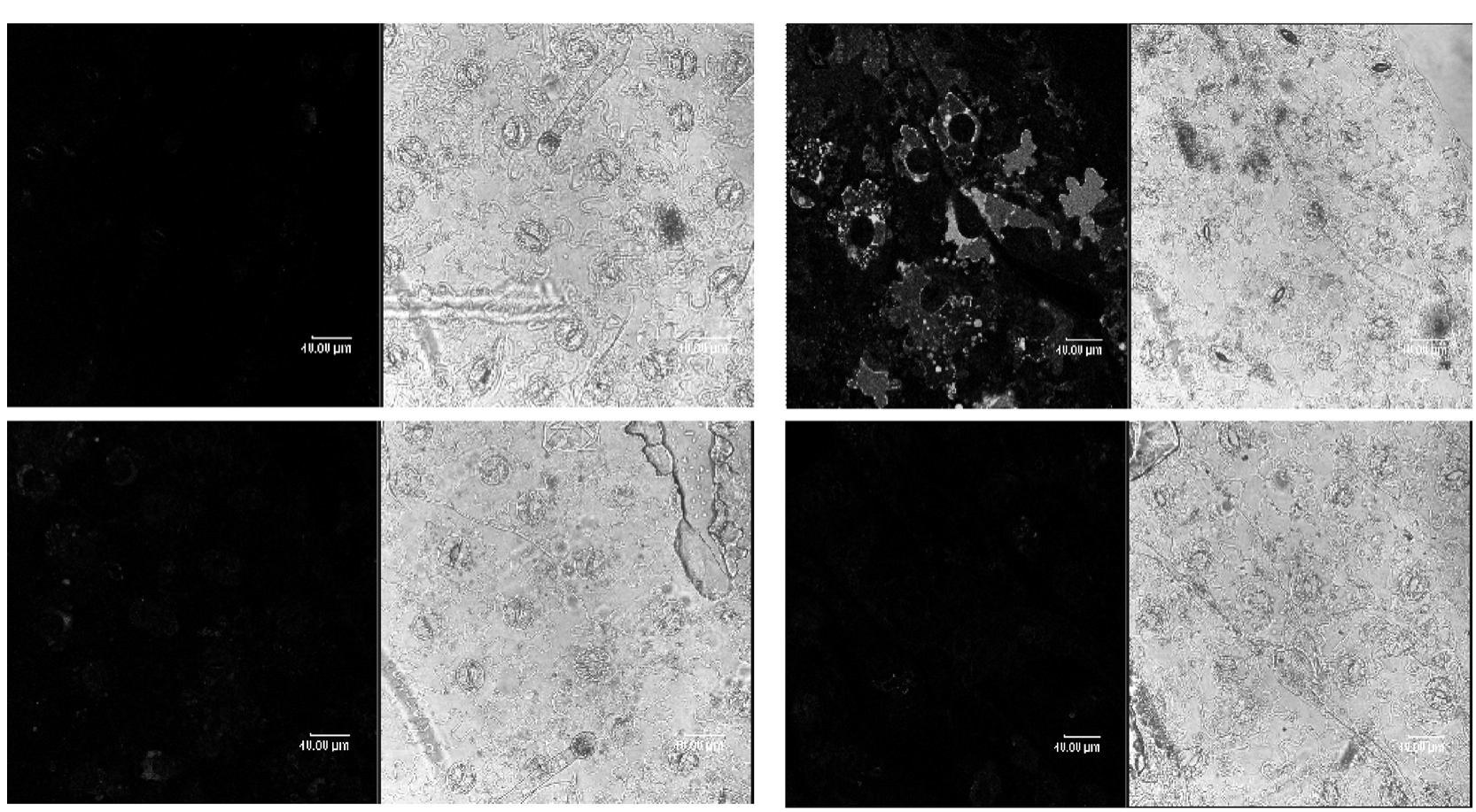
Because of activity of SOD and CAT induced by OGAs and the two enzymes correlative with oxygen free radical,we examined the production of H2O2 induced by OGAs.To study the effects of OGAs on the production of H2O2 in tobacco cells,the H2O2-sensitive fluorophore H2DCF-DA were used.The results of production of H2O2 in epidermal cells of tobacco leaves induced by OGAs were shown in Figure 4.It was found that OGAs caused an increase of intracellular H2DCF-DA fluorescence in epidermal cells and guard cells of tobacco leaves,indicating the production of H2O2.Fluorescence became visible along the plasma membrane and in organelles in the epidermal cells of tobacco leaves treated with OGAs(Figure 4C),but the fluorescence was very faint in the epidermal cells only loaded with H2DCF-DA(Figure 4A).The Figure 4E and G showed that CAT and DPI could inhibit the level of H2DCF-DA fluorescence in the cells of tobacco leaves treated with OGAs.The results revealed that CAT and DPI could suppress the production of H2O2.
Figure 4 Laser scanning confocal microscopy of OGA-induced production of H2O2 in epidermal cells of tobacco leaves. (A) The cells loaded with H2DCF-DA. (B) Bright field image of the cells loaded with H2DCF-DA. (C) The cells loaded with H2DCF-DA before treatment with OGA. (D)Bright field image of the cells loaded with H2DCF-DA before treatment with OGA. (E) The cells loaded with H2DCF-DA and elicited by OGA in the presence of the CAT. (F) Bright field image of the cells loaded with H2DCF-DA and elicited by OGA in the presence of the CAT. (G) The cells loaded with H2DCF-DA and elicited by OGA in the presence of the DPI. (H) Bright field image of the cells loaded with H2DCF-DA and elicited by OGA in the presence of the DPI.
The NO-sensitive fluorophore DAF-2DA was used to observe NO accumulation.The observed LSCM results of OGAs-induced production of NO in epidermal cells of tobacco leaves were shown in Figure 5.It was found that OGAs could enhance the level of intracellular DAF-2DA fluorescence in epidermal cells of tobacco leaves,indicating massive production of NO.Production of NO and/or accumulation was observed in organelles and along the plasma membrane in the epidermal cells of tobacco leaves treated with OGAs(Figure 5C).However,the DAF-2DA fluorescence indicating production of NO was not observed in the epidermal cells only loaded with DAF-2DA(Figure 5A).The results also indicated that CPTIO and L-NAME could inhibit the level of H2DCF-DA fluorescence in the cells of tobacco leaves treated with OGAs(Figure 5E and G).The results representedthat CPTIO and L-NAME could suppress the production of NO.
Figure 5 Laser scanning confocal microscopy of OGA-induced production of NO in epidermal cells of tobacco leaves. (A) The cells loaded with DAF-2 DA. (B) Bright field image of the cells loaded with DAF-2 DA. (C) The cells loaded with DAF-2 DA before treatment with OGA. (D) Bright field image of the cells loaded with DAF-2 DA before treatment with OGA. (E) The cells loaded with DAF-2DA and elicited by OGA in the presence of the CPTIO. (F) Bright field image of the cells loaded with DAF-2DA and elicited by OGA in the presence of the CPTIO. (G) The cells loaded with DAF-2DA and elicited by OGA in the presence of the L-NAME. (H) Bright field image of the cells loaded with DAF-2DA and elicited by OGA in the presence of the L-NAME.
3.5 Assay for the effect of OGAs and exogenous NO and H2O2 on disease symptom
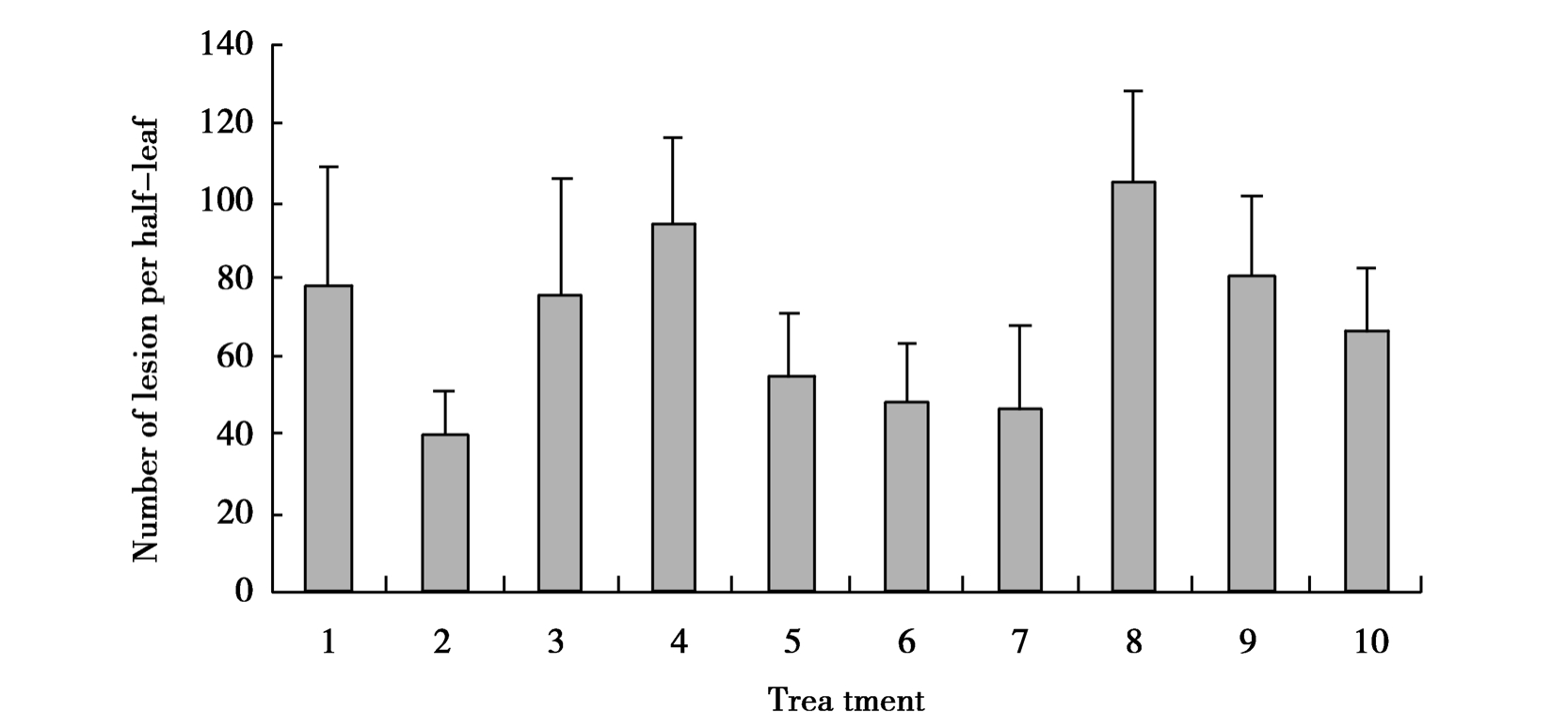
As H2O2 and NO appear to be a key factor associated with plant induced defense disease,it was interesting to test the effect of exogenous NO and H2O2.The effect of OGAs,NO donor SNP and H2O2 at different concentrations and some scavengers are summarized in Figure 6.It was found that treatment with OGAs,SNP and H2O2 protected tobacco leaves against TMV local infection.The least lesion was observed at the treatment of 50μg/ml OGAs among the all treatments.The inhibition effect of H2O2 showed dependence on the amount of H2O2.The lesion of co-treatment of OGAs and the H2O2 scavenger CAT or ascorbic acid on TMV infection was as high as CK.We also observed SNP inducing resistance was dose-dependent.When the tobacco plants were treated with L-NAME before OGAs,the induced resistance was depressed.Therefore,we can presume NO and H2O2 are important factors participating in OGAs inducing resistance to TMV.
Figure 6 Effect of OGAs and exogenous NO and H2O2 on disease symptom
4 Discussion
Pectic oligosaccharides,produced by microbial enzymes,are well-known oligosaccharins,eliciting defence responses in diseased plants(Dumville et al.,2000).A broad spectrum of OG-induced pathogenesis-related defense responses has been reported(M.T.Esquerré-Tugayé et al.,2000).Most defense and developmental responses are induced by OGAs with a degree of polymerization(DP)between 10 and 15 galacturonic acid residues.OGAs with a DP less than 8 can also trigger defense responses in plants:they induce accumulation of protease inhibitors(T.Moloshok et al.,1992),ethylene production(S.D.Simpson et al.,1998)and elicitation of genes involved in jasmonic acid metabolism in tomato(C.Norman et al.,1999).In this report,we observed the OGAs with a DP between 2~8 could induce tobacco resistance to TMV.The concentration of OGAs used was also discussed.OGAs-induced plant growth has been reported(LoSchiavo et al.,1991;Filippini et al.,1992),and the maximal effect to growth was about 10-4 M(Stephen et al.,1993).To elicit plant defense responses,OGAs concentration higher than those usually required for control developmental process.In our experiments,50μg/ml was the best concentration to induce resistance within 100μg/ml(data not shown).It showed the efficiency of the OGAs in inhibition of virus infection was not depended on the dose of OGAs.But the inhibition effect was dependent on the treatment time.We observed the inducing effect of resistance to TMV was gradually elevated before 19d,but the mechanism of this needed further study.Research showed that lag period of the induced resistance of glucohexaose was about 7days and the protection period was about 28 days(Li Hongxia et al.,2005).Furthermore,tobacco plants treated by sulfated fucan or linear β-1,3 glucan showed resistance to TMV or bacterium E.carotovora after 5 days(Olivier Klarzynski et al.,2003;2000).So far no oligosaccharides were reported to have so long time inducing effect.Therefore,OGAs have more predominance to be applied in agriculture.
Experimental results also showed that NO and H2O2 played important roles in OGAs inducing tobacco resistance to TMV.NO and H2O2 as important signaling active molecules in pathogen defense reaction has been extensively studied(Levine et al.,1994;Mehdy et al.,1996;Baker et al.,1995;Jabs et al.,1996;Delledonne et al.,1998;Rout-Mayer et al.,1997?;Binet et al.,1998).First,we examine the activity of plant resistance correlated enzymes.Because the activity of PAL has been confirmed elevated by many reports(Messiaen et al.,1994;Lapous et al.,1998;Dixon et al.,1989;Tepper et al.,1990),we just mensurated the PPO,POD,SOD and CAT.This includes the activity of SOD and CAT elevated,so we estimated the extra H2O2 production.
To evaluate the stimulatory effect of OGAs on tobacco cells,we measured the production of H2O2 and NO in tobacco cells.The data indicated that OGAs induced the production of H2O2 and NO in epidermal cells of tobacco within a short time.These results were in agreement with the reports by Xiangyang Hu,who claimed OGAs stimulated NO accumulation in the growth medium of ginseng suspension cultures(Hu et al.,2003).Rout-Mayer and Binet discovered respectively H2O2 production within a few minutes after the addition of OGAs to suspension-cultured tobacco cells(Rout-Mayer et al.,1997;Binet et al.,1998).
Many reports show H2O2 and NO exist are correlated to plant defense.H2O2 is involved in the induction and/or execution of hypersensitive reaction(C.S.Bestwick et al.,1997).H2O2 is required for the cross-linking of plant cell wall components as a part of the structural defense response(C.Lamb et al.,1997).The production of H2O2 may also lead to the development of an antimicrobial environment within the apoplast(M.Peng et al.,1992).In many cases,H2O2 collaborate with NO to execute invading pathogens.H2O2 and NO production were induced almost at the same time by cryptogein,a fungal elicitor(Foissner et al.,2000).NOS inhibitors compromise the hypersensitive resistance response in Arabidposis and tobacco(Delledonne et al.,1998?;Huang et al.,1998).TMV infection could elevate NOS(nitric oxide synthase)activity,and NO could induce PR-1 expression(Durner et al.,1998).NO,as well as other ROS,have been shown to stimulate the accumulation of SA(Durner et al.,1999),which play a critical signaling role in the activation of plant defense responses after pathogen attack.
Furthermore,to test whether OGAs functions on inducing resistance in tobacco via NO and H2O2 pathway,we examined the effects of OGAs,exogenous NO donor SNP and H2O2 on inducing resistance to TMV.It was found that all of these treatments reduced lesion caused by TMV.But co-treatment with OGAs and H2O2 scavenger CAT or ascorbic acid blocked the inducing resistance.The tobacco plants inhibited NOS activity by L-NAME were not induced resistance by OGAs.So the defense response induced by OGAs was connected with NO and H2O2 pathway.
The study reported herein reveals that OGAs can induce the production of H2O2 and NO,and induce the defense response against TMV.Our understanding of OGAs induced resistance is sketchy.The mechanisms of OGAs eliciting defense responses of tobacco need further investigation.