Function of Predatory Nematode and Plant-parasites Nematode in Different Tillage Practices*
Function of Predatory Nematode and Plant-parasites Nematode in Different Tillage Practices*
土壤线虫具有丰富的生物多样性。多数土壤线虫生态学者将土壤线虫按照取食习性分成植物寄生线虫、食细菌类线虫、食真菌类线虫、捕食类或杂食类线虫、以基质为食的线虫和取食单细胞的线虫等
捕食性线虫属于自由生活线虫,通常生活周期长、繁殖能力低和对环境干扰敏感。一般不是土壤中的优势种群,但能更好地指示土壤状况的较长期变化
1 材料与方法
1.1 试验材料
供试作物:大豆(
1.2 试验方法
丰度(Abundance)=100g干土中土壤线虫的数量
相对丰度(Relative Abundance)RA=丰度/线虫总数
2 结果与分析
2.1 线虫种群的多样性
(1)孔咽属线虫
分类地位:矛线目,矛线亚目,矛线总科,矛线科。
形态特征:食道前部细长,后部扩大呈圆筒形或梨形,口腔内常有1个轴针或壁齿。虫体大,通常大于4mm。体表由微弱棕纹和交织纹,偶尔也有横环纹。唇区突出,唇片分离。齿针粗,开口大于齿针1/2。侧器由中央支撑。食道在中部扩大。背食道腺与开口间距离大。有贲门盘。雌虫卵巢1对,阴门横裂。雄虫有矛线形交合刺,有侧引带,中腹生殖乳突数个或多个。尾短,圆锥形或半球形,雌雄尾形相似。
(2)前矛线属
分类地位:矛线目,矛线亚目,矛线总科,矛线科。
形态特征:口腔内常有一个轴针或壁齿,口腔壁无角质化或肋围绕着轴针,食道前部细小,后部扩大成圆筒形或梨形,食道后部扩大部等于或大于1/3食道长,口针较短,延伸部简单,无凸缘或基部球,咽具有一轴针,口针周围无角化板,食道前部正常肌肉质,唇区不开阔,口针导环明显,无导鞘,内环乳突不在头部外侧,两性尾部皆为丝状。
(3)锉齿属线虫
分类地位:单齿目,锉齿科,锉齿属。
形态特征:食道呈圆筒形,基部稍宽,口腔壁角化,至少有1个粗大的背齿。背齿位于口腔前部向前指,相对位置上有2列边缘齿。经温和热杀死后向腹面弯曲,唇区分开,口囊腹壁小齿6排,雌虫双卵巢,尾短,圆锥形,在尾近中部突然向腹面弯曲。
(4)单齿属线虫
分类地位:单齿目,单齿科。
形态特征:虫体长1~6mm。食道呈圆筒形,基部稍宽,口腔壁角化,至少有1个粗大的背齿。口腔卵圆形。背齿大,位于口腔前部1/3,齿尖向前,无亚腹齿。背齿相对处亚腹侧口腔壁有2道横肋。卵巢1对,偶单个。雄虫常见,交合刺细长。生殖附器10~47个不等。雌雄尾形相似。尾腺和尾腺管明显。
在分离鉴定的4个属线虫中,孔咽属在土壤线虫总数中占优势,属于亚优势属。在连作田和撂荒田中仅发现了孔咽属;在水改旱中捕食性线虫的种类最多,4个属的线虫均有出现,其中孔咽属和锉齿属为常见属,前矛线属和单齿属为一般属。轮作中没有发现单齿属线虫,而其他3属线虫均存在,但孔咽属明显少于其他3种耕作方式。不同耕作方式大豆田中,捕食性线虫的种类和数量均有差异(表1)。
OrderGeneraContinuouscroppingRotationPaddy-uplandFallowDorylaimidaAporcelaimus9.51%b1.40%c4.48%c6.84%bProdorylaimus0.01%d0.27%dMononchidaMylonchulus0.02%d1.10%cMononchus0.07%d
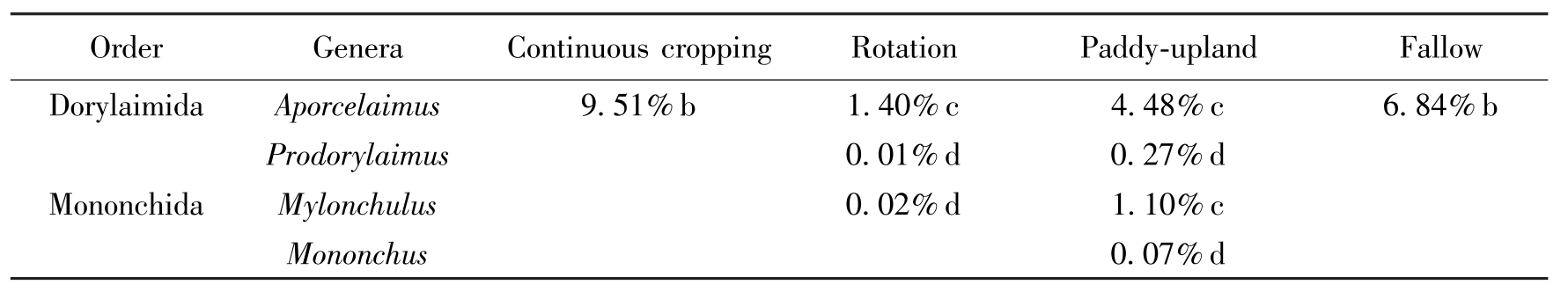
Table 1 The genus, relative abundance and dominance of predatory nematode in four tillage practice
OrderGeneraContinuouscroppingRotationPaddy-uplandFallowTylenchidaHelicotylenchus30.42%a90.55%a71.45%a41.53%aPararotylenchus0.04%d—0.01%d0.17%dScutellonema0.09%d——Criconemella—0.01%d——Hoplolaimus0.01%d0.01%d——Pratylenchus—0.42%d——Paratylenchus0.06%d0.01%d—0.07%dHeterodera35.47%a1.09%c0.14%d26.34%aXiphinema0.15%d——Neothada—0.01%d—0.01%dTylenchus0.19%d—0.54%d—Filenchus0.37%d0.53%d0.11%d—Psilenchus0.13%d0.43%d0.50%d—Boleodorus0.07%d—1.07%c—Basiria0.19%d0.01%d0.16%d0.30%d
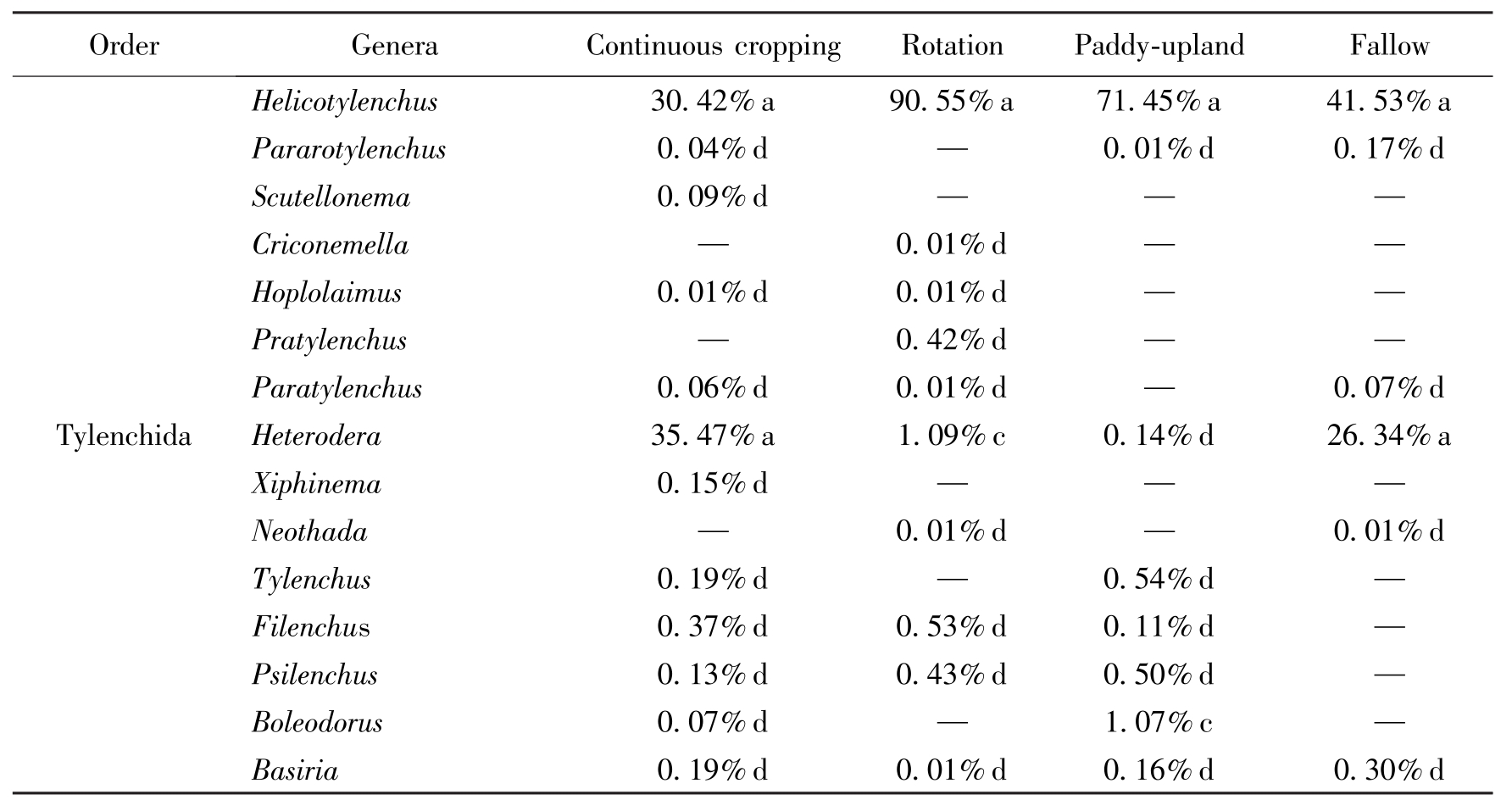
Table 2 The genus, relative abundance and dominance of plantparasitic nematodes in four tillage practice
2.2 大豆不同生育时期植物寄生线虫和捕食性线虫的总量变化
试验结果显示,随着大豆生育期的变化,不同耕作方式下大豆田植物寄生线虫变化基本一致,均是在大豆播种期、苗期和开花期线虫的数量变化有逐渐减少的趋势,到结荚期线虫的数量明显增多,成熟期达最大。轮作中植物线虫总量远高于连作、水改旱和撂荒,变化最明显,其次是水改旱和撂荒。在4种耕作方式中,连作的每个时期(除播种期外)植物线虫数量都为最少,到成熟期数量增长也不显著,变化最平稳。
大豆田中捕食性线虫数量比植物寄生线虫的数量少近50倍,不是土壤中的优势种群,但对环境变化较为敏感。不同耕作方式下捕食性线虫随大豆生育时期变化的趋向不同。连作和轮作中捕食性线虫变化的曲线大致相同,从播种期到成熟期先减少后增多。水改旱地块中前3个时期捕食性线虫的数量几乎没有变化,结荚期和成熟期急剧增加达到群体数量最大。撂荒地中捕食性线虫的数量在播种期最多,而到苗期、开花期则减少到几乎没有,结荚期时的数量又上升到4种耕作方式中的最高值,到成熟期又有所降低(图1)。
2.3 大豆不同生育期捕食性线虫和植物寄生线虫数量关系
不同生育期植物寄生线虫和捕食性线虫的变化曲线基本一致,从播种期到苗期、开花期缓慢减少,从开花期到成熟期逐渐增多,成熟期达到最大(图3)。从图中曲线可看出捕食性线虫的变化较植物寄生线虫有些迟滞,说明捕食性线虫的数量有可能是随着植物寄生线虫数量的增减而变化,这与土壤中线虫食物链有一定的关联性。
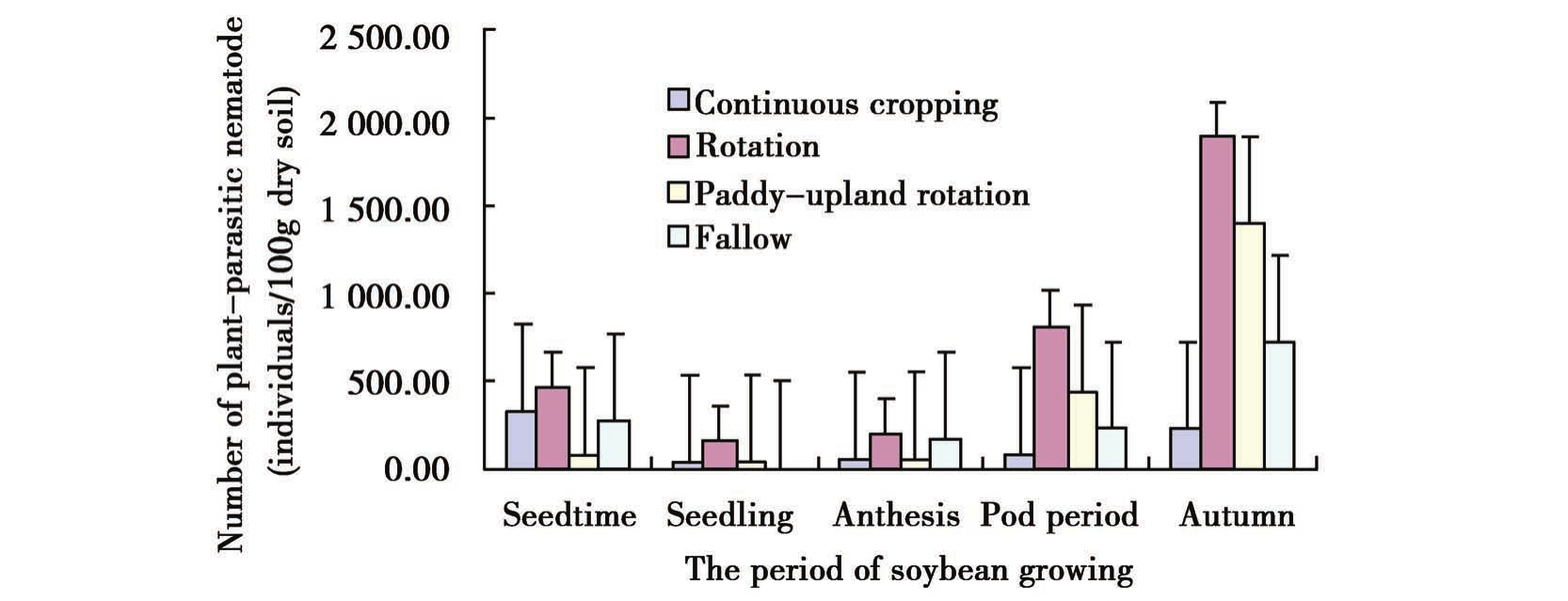
Figure 1 Changes in numbers of plant-parasitic nematode and predatory nematode during soybean period in different tillage practices
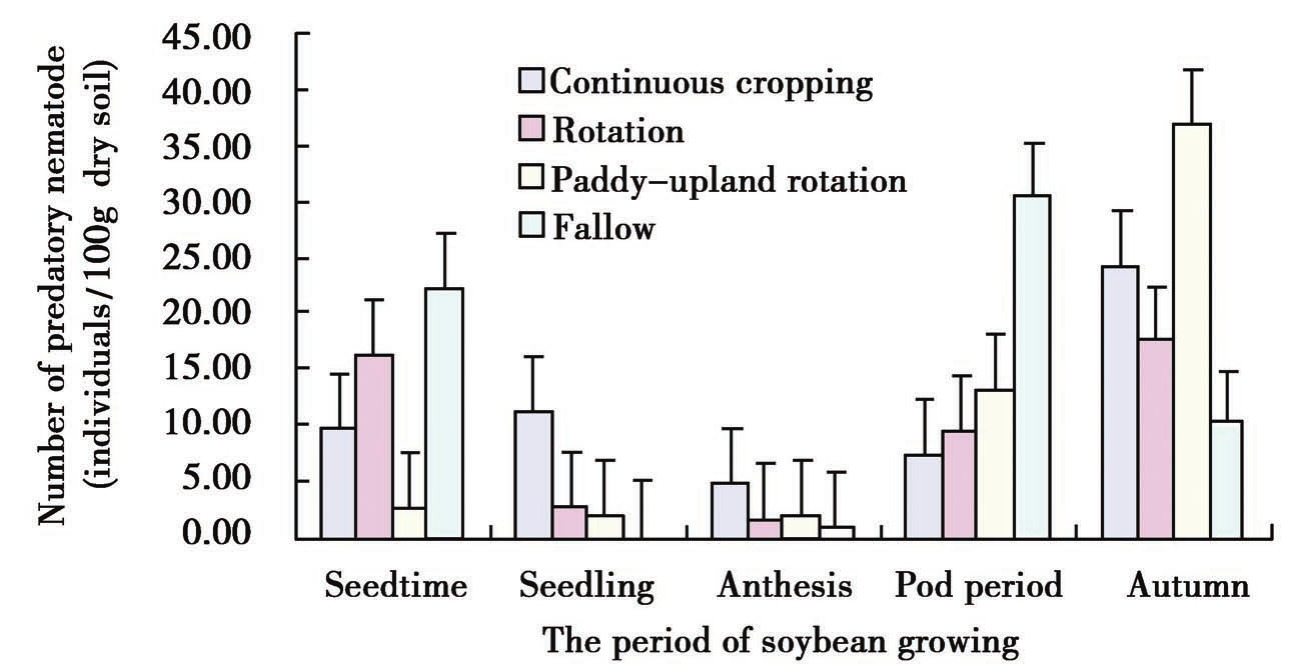
Figure 2
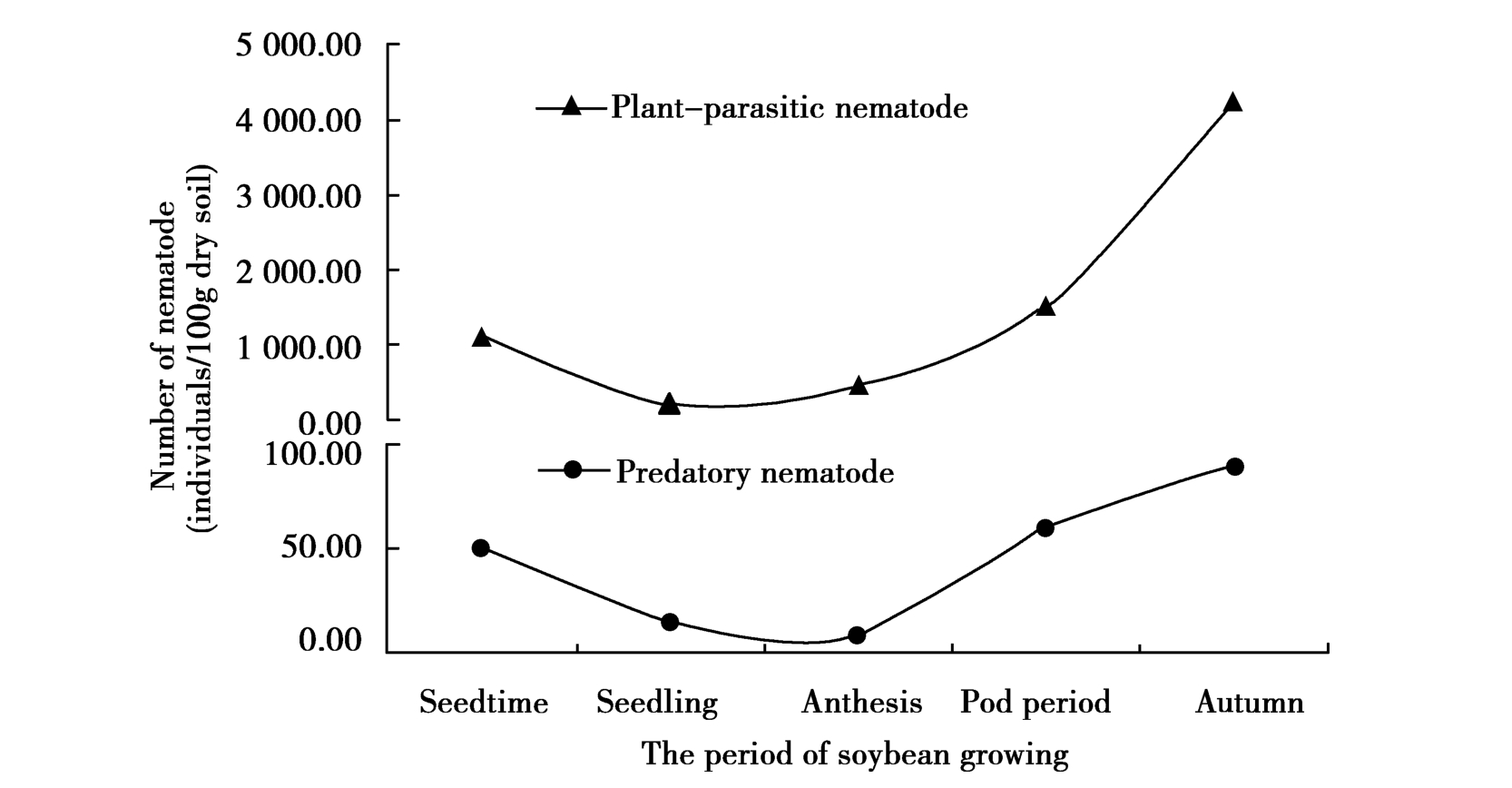
Figure 3 Dynamic change of plant-parasitic nematode and predatory nematode in soybean period
3 结论与讨论
本研究仅在大豆田发现了4属的捕食性线虫,其中孔咽属为亚优势属,存在于4种耕作方式中,而一般属锉齿属和单齿属仅存在于水改旱和轮作中;植物线虫类群中螺旋属在4种耕作方式中均为优势属,胞囊属在连作和撂荒中为优势属,在轮作和水改旱中则属于一般属;但植物线虫总数在轮作中最多,水改旱其次,撂荒和连作最少。
在农田生态系统中,捕食性线虫的生育周期长而且繁殖速度慢,受到人为扰动后群体数量恢复较慢,因此该类线虫中的某些种类可以作为生物指示剂指示土壤的健康情况。本研究同一种作物在不同的耕作方式下,植物线虫和捕食性线虫的种群多样性主要体现在种属分布不同,数量的多寡不同。在水改旱和轮作中的捕食性线虫的种类比连作和撂荒中的丰富,而且胞囊线虫的相对丰度也很低,仅在1%左右。胞囊线虫作为大豆的重要病原物起着指示大豆胞囊线虫病害发生严重度的作用。因此,研究认为,植物线虫类群中的胞囊属和捕食性线虫类群中的锉齿属和单齿属对土壤的扰动较为敏感,能很好地反映土壤发病和健康状况的变化,可作为土壤中重要的指示生物。
本文的研究显示在大豆不同生育期,捕食性线虫的种群数量随着植物寄生线虫的增加而增加。植物寄生线虫会追随植物的生长发育而大量繁殖自身,因此在植物生育末期种群数量一般会增殖到最大,而捕食性线虫以土壤中线虫或其他生物为食,因此也会随植物线虫群体数量的增大而增加。Das等人通过对耕作田和草坪土壤中植物寄生线虫、捕食性线虫和自由生活线虫种群密度动态变化的研究中发现捕食性线虫的动态变化依赖于被捕食线虫种群多样性和密度









