Effects of Secondary Metabolites of Actinomycetes on Secondary Juvenile Mortality of Heterodera Glycines*
Effects of Secondary Metabolites of Actinomycetes on Secondary Juvenile Mortality of Heterodera Glycines*
大豆胞囊线虫,
放线菌作为抗生素的主要产生菌,其代谢途径复杂,产生的次生代谢产物在结构类型和生物活性等方面都呈现出与细菌和真菌不同的特点和多样性
大豆胞囊线虫作为一种专性寄生物,存在明显的生理分化现象。一个线虫田间群体不是寄生性整齐一致的单一小种,连续种植抗病品种,由于抗病基因对线虫群体的选择作用,田间线虫的生理小种类型会发生改变
1 材料与方法
1.1 供试菌株
本试验采用的菌株C25-3分离自吉林省辽源市,H-4和H-2分离自黑龙江省哈尔滨市延寿县,C44和C49分离自辽宁省沈阳市。
1.2 大豆胞囊线虫3号生理小种胞囊的分离和二龄幼虫的孵化
1.3 田间不同群体的大豆胞囊线虫胞囊和二龄幼虫的获得
从黑龙江省农业科学院大庆分院安达盐碱地作物研究所大豆育种基地随机五点取样,采用改良淘洗—过筛法从土壤中分离胞囊。孵化方法同1.2.2。
1.4 放线菌菌株代谢物对靶标线虫的作用
取制备好10
1.5 计算方法
线虫死亡率(%)=死亡线虫数/供试线虫数×100%
校正死亡率(%)=(处理线虫死亡率-对照线虫死亡率)/(1-对照线虫死亡率)×100%
2 结果与分析
2.1 放线菌菌株代谢物对SCN3号生理小种新鲜胞囊孵化J2的影响
28℃条件下振荡培养7天得到的发酵液经离心后,在4×稀释浓度下,24h对大豆胞囊线虫3号生理小种新鲜胞囊孵化的二龄幼虫均有一定的致死作用。试验结果表明,C25-3处理的二龄幼虫校正死亡率达到95.23%,活性最高,与其他菌株相比差异均显著。H-2、C49和H-4对二龄幼虫校正死亡率相近,均达到70%以上,C44对二龄幼虫的校正死亡率为62.80%,活性最低,但均与对照相比达极显著差异水平(表1)。
StrainsMortalityofjuvenile(%)ⅠⅡⅢAverage(%)Correctedratio(%)C25-3100.093.3392.8695.40aA95.23H-477.7869.2372.7373.25bcBC72.30H-286.3680.0071.4379.26bB78.53C4981.2571.4378.5777.08bBC76.27C4455.5670.0066.6764.07cC62.80CK5.005.260.003.42dD—
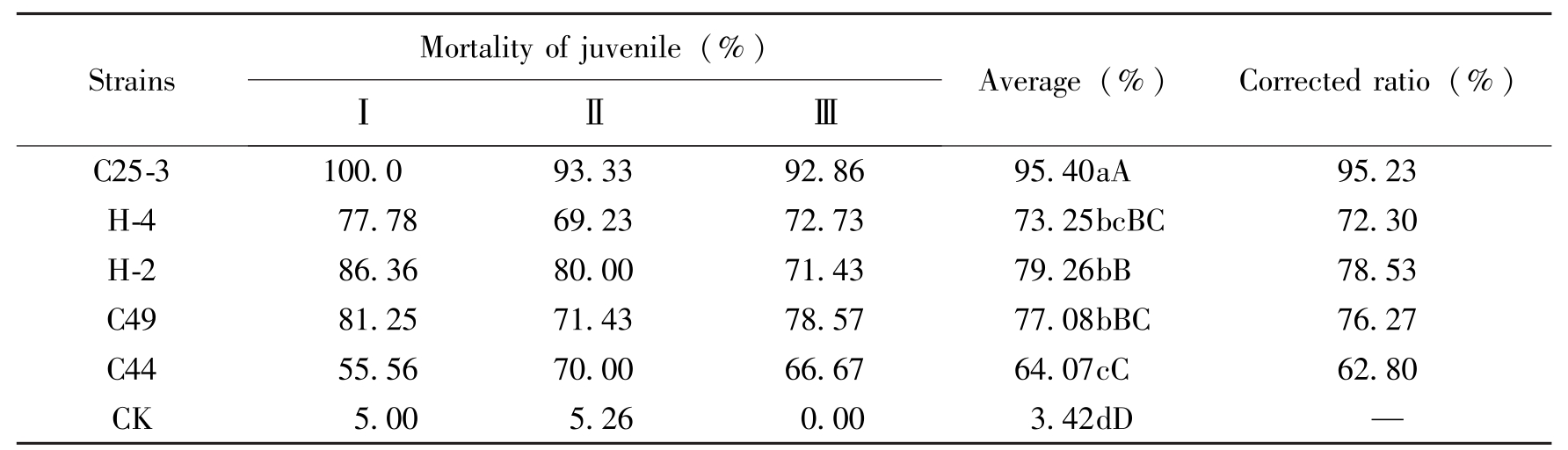
Table 1 The effect of different strains to SCN J2 of young cyst of race 3
2.2 放线菌菌株代谢物对SCN3号生理小种越冬胞囊孵化J2的影响
试验结果表明,C25-3对大豆胞囊线虫3号生理小种越冬胞囊孵化的二龄幼虫有较高的毒杀作用,发酵液稀释4倍处理24h后,二龄幼虫校正死亡率达到94.84%,与其他几株相比差异均极显著。C49、C44、H-4和H-2对处理的二龄幼虫校正死亡率差异不显著,均在70%左右。5株菌与对照相比均达极显著差异水平(表2)。
StrainsMortalityofjuvenile(%)ⅠⅡⅢAverage(%)Correctedratio(%)C25-3100.0091.6792.8694.84aA94.84H-479.1760.0070.0069.72bB69.72H-264.2986.6762.5071.15bB71.15C4970.0072.7370.0070.91bB70.91C4455.5675.0077.7869.45bB69.45CK0.000.000.000.00cC—
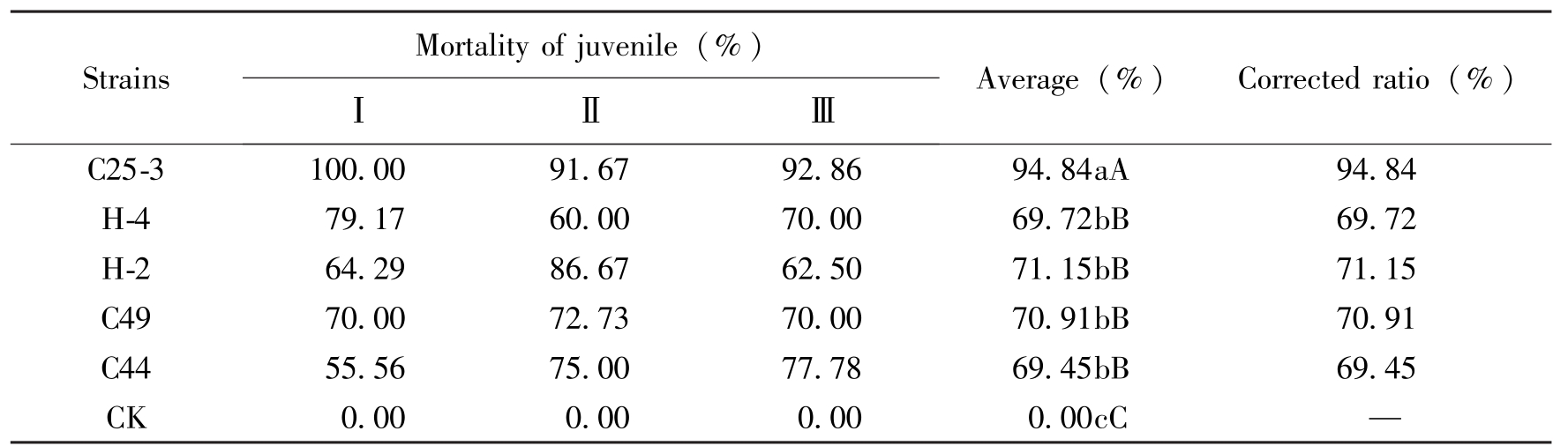
Table 2 The effect of different strains to SCN J2 of old cyst of race 3
2.3 放线菌菌株代谢物对田间混合群体的大豆胞囊线虫J2的影响
试验结果表明,5株菌对田间混合群体的大豆胞囊线虫J2均有不同程度的抑制作用,与对照相比达极显著差异水平。其中C25-3处理后J2的校正死亡率达96.59%,与其他几株相比差异均极显著。H-4对二龄幼虫的活性次之,校正死亡率为87.29%。C44和H-2为中等水平,
StrainsMortalityofjuvenile(%)ⅠⅡⅢAverage(%)Correctedratio(%)C25-3100.0090.00100.0096.67aA96.59H-472.73100.0090.0087.58aAB87.29H-280.0080.0063.6474.55bBC73.97C4470.0054.5570.0064.85bcCD64.05C4975.0063.6472.7354.17cD53.13CK0.000.006.672.22dE—
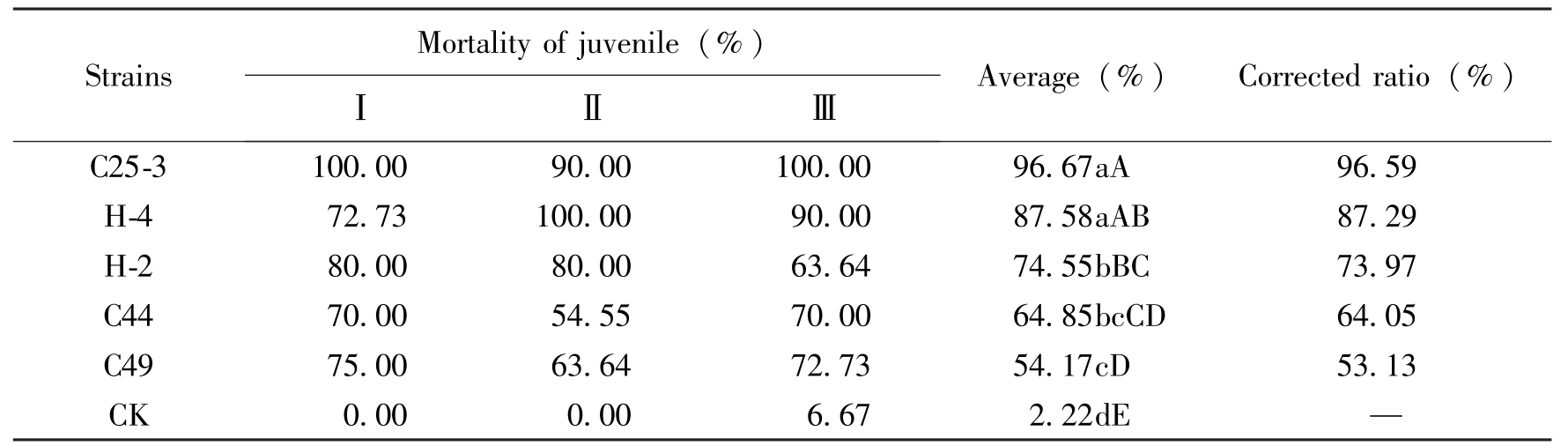
Table 3 The effect of different strains to SCN J2 of different races
3 结论与讨论
本研究结果显示,筛选出的5株放线菌菌株中,C25-3对J2的活性最高,另外,C25-3,H-2和C44对大豆胞囊线虫J2的毒性不因线虫的致病性和活性差异而改变,而C49和H-4表现出因线虫的致病性和活性差异而改变的特性。目前,人们正在探索如何筛选高效、稳定的线虫生防放线菌的方法。Skantar根据胞囊孵化时间的快慢将大豆胞囊线虫的胞囊分为TN17和TN18两种类型,室内研究了格兰德霉素(Geldanamycin)对TN17和TN18孵化的二龄幼虫活性的影响。结果表明:格兰德霉素对来源不同的胞囊孵化的二龄幼虫活性有一定的差异
本文研究的不同放线菌次生代谢产物对胞囊混合群体孵化出的二龄幼虫毒杀活性表现出差异,说明生防菌对J2的作用机制不同,这可能是由于大豆胞囊线虫的生理分化而导致二龄幼虫的抗性存在差异;还有可能是放线菌的次生代谢产物复杂,往往含有多个有效组分,导致对不同胞囊来源的二龄幼虫的活性存在差异。
生防放线菌是一种很有潜力的生物防治资源,对于活性菌株的次生代谢产物中活性物质的分离纯化以及安全性评价还有待进一步研究。生防放线菌除产生抗生素之外,还可能同时具有多种生防机制,必须全面分析和利用,才能达到最佳的防治效果,从而控制大豆胞囊线虫的为害。









