Review of the Mechanism of Action and Resistance of Systemic Fungicides used for Controlling Oomycete Disease
Review of the Mechanism of Action and Resistance of Systemic Fungicides used for Controlling Oomycete Disease
Cavalier-Smith(1981;1988)提出将细胞生物分为八界,《真菌词典》第八版(1995)和第九版(2001)均接受了这一分类系统。在这一分界系统中,卵菌门(Oomycota)属于色菌界(Chromista),有1纲9目及其他地位待定的4目,其中引起植物病害的有水霉目(Saprolegniales)、腐霉目(Pythiales)、指梗霉目(Sclerosporales)和霜霉目(Peronosporales)
在对卵菌的综合防治的措施中,化学防治仍然是控制卵菌病害的主要手段之一,但目前生产中普遍发生了卵菌对现有杀菌剂的抗药性问题,这对当前农业的发展构成了严重的威胁
病害名称病原物病害名称病原物十字花科白锈病白锈菌(Albugocandida)马铃薯晚疫病致病疫霉菌(Phytophthorainfes-tans)谷子白发病禾指梗霉菌(Sclerosporagra-minicola)瓜类疫病掘氏疫霉菌(P.drechsleri)烟草猝倒病瓜果腐霉菌(Pythiumaphanider-matum)辣椒疫病辣椒疫霉菌(P.capsici)黄瓜霜霉病古巴假霜霉菌(Pseudoperonospo-racubensis)柑橘褐腐柑橘褐腐疫霉菌(P.citrophthora)葡萄霜霉病葡萄生轴霜霉菌(Plasmoparaviticola)
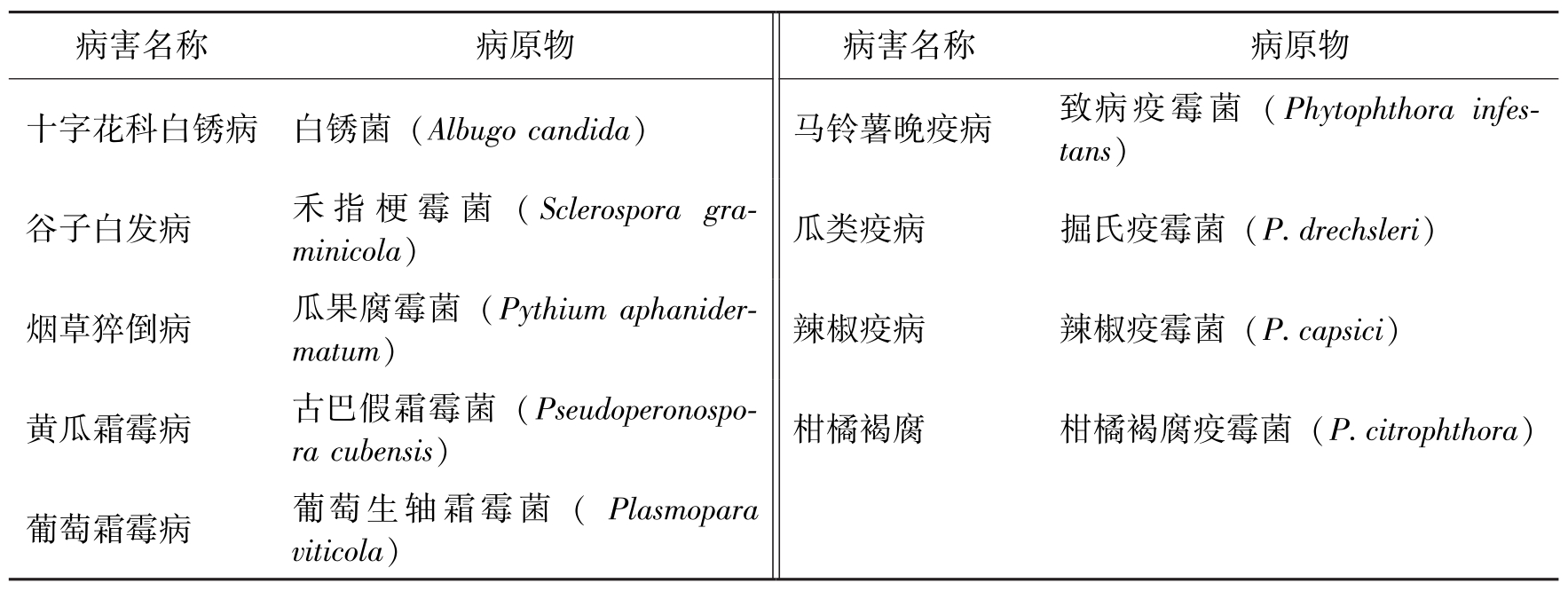
表1 我国卵菌引起的主要植物病害
1 苯基酰胺类杀菌剂
苯基酰胺类杀菌剂的特点是低毒,选择性强,可以被植物的根、叶和幼茎迅速吸收,并通过导管和细胞间隙等质外体系统向植株上部转移
Dr.P.A.Urech在1977年研究开发了甲霜灵,它是苯基酰胺类杀菌剂中第一个商品化的品种,其特点是病菌在植物体内形成吸胞后抑菌作用才能发挥,对卵菌侵入寄主前的各阶段,如游动孢子的释放、萌发和侵入的抑制作用不明显,但对侵入寄主后的各阶段,如菌丝在植物体内的生长、吸器形成及孢子囊的产生等有显著的抑制作用
最初对苯基酰胺类杀菌剂容易引起病原菌抗药性的问题并未引起注意,直到 1981年荷兰的Dekker发现甲霜灵单剂在葡萄上使用仅一个季度就发生田间抗药性之后才给以重视。Davides等
通过对抗药和敏感菌株杂交后代的分析,表明抗药性是由隐性的单基因或核内的单作用位点控制,发展速度快,抗性水平高,抗药菌株遗传稳定,适合度、致病力几乎与野生的敏感菌相同,故经过施药的选择敏感菌很快消亡,抗性菌在群体中得以繁殖发展,并很快成为优势种群,出现田间突然药剂失效
2 丙烯酰胺类杀菌剂
丙烯酰胺类杀菌剂中对卵菌病害有很好防治效果的药剂主要有烯酰吗啉和氟吗啉,其作
烯酰吗啉在1988年由Shen Group公司研制开发。烯酰吗啉可强烈抑制游动孢子囊的形成、休眠孢子的萌发和菌丝生长,但不影响游动孢子的释放。与菌丝细胞壁合成相比,孢子囊壁的合成对烯酰吗啉更为敏感。Bartnicki
烯酰吗啉1992年获得注册登记并投放市场,尽管已经使用多年,但国内外的室内和田间实践均证明,烯酰吗啉在田间表现为低抗药性风险。Dereviagina等
3 甲氧基丙烯酸酯类杀菌剂
甲氧基丙烯酸酯类杀菌剂是以天然抗生素Strobilurin A为先导化合物而开发的新型杀菌剂,自1969年Musikek及其合作者首先报道以来,对此类化合物的研究已成为杀菌剂开发的新热点。这类杀菌剂具有杀菌谱广和杀菌活性高的特性,具有保护、治疗、铲除、渗透作用,对环境和非靶标生物安全
甲氧基丙烯酸酯类杀菌剂的作用位点独特,不同于以往任何抑制剂作用于呼吸链上的位置。Cyt bc1呼吸抑制剂有两类:一类是位于线粒体内膜内壁Qi位点(CoQ的还原位点)的cyt b高势能血红色素结合的抑制剂,这类抑制剂称为Qi位点抑制剂,简称QiIs,如抗霉素,cyanoimidazole等。另一类与位于线粒体内膜外壁的Qo位点(CoQ的氧化位点)的cyt b低势能血红色素结合的抑制剂,这类抑制剂称为Qo位点抑制剂,简称QoIs,如甲氧基丙烯酸酯类杀菌剂、咪唑菌酮、 唑菌酮等
唑菌酮等
甲氧基丙烯酸酯类杀菌剂为能量生成抑制剂,其作用机理是通过与真菌细胞色素b上的辅酶Q
在甲氧基丙烯酸酯类杀菌剂开发和应用的早期,人们认为这类杀菌剂的抗性风险是中等水平,但是1998年德国使用苯氧菌酯防治小麦白粉病2年后发现防效明显下降,原因是小
4417407(咪)唑类杀菌剂
 (咪)唑类杀菌剂活性高、作用机理独特,与现有杀菌剂无交互抗性,能与多种杀菌剂混配,可有效缓解内吸性杀菌剂的抗性问题。
(咪)唑类杀菌剂活性高、作用机理独特,与现有杀菌剂无交互抗性,能与多种杀菌剂混配,可有效缓解内吸性杀菌剂的抗性问题。
 唑菌酮由杜邦公司开发,对锈病、霜霉病、晚疫病等均具有良好保护、治疗、渗透、内吸等作用。它对病原菌在生长过程中所释放出的孢子影响最大,药剂一旦与孢子接触,孢子即停止活动,出现崩死。另外,对孢子的萌发和菌丝的伸长也有一定的抑制作用
唑菌酮由杜邦公司开发,对锈病、霜霉病、晚疫病等均具有良好保护、治疗、渗透、内吸等作用。它对病原菌在生长过程中所释放出的孢子影响最大,药剂一旦与孢子接触,孢子即停止活动,出现崩死。另外,对孢子的萌发和菌丝的伸长也有一定的抑制作用
 唑菌酮能抑制病原菌线粒体的电子传递,主要阻断细胞色素b和细胞色素c之间的电子传递通道中ADP-ATP的氧化磷酸化作用,使病原菌无法产生所必需的能量。
唑菌酮能抑制病原菌线粒体的电子传递,主要阻断细胞色素b和细胞色素c之间的电子传递通道中ADP-ATP的氧化磷酸化作用,使病原菌无法产生所必需的能量。
 唑菌酮能抑制病原菌线粒体的电子传递,主要阻断细胞色素b和细胞色素c之间的电子传递通道中ADP-ATP的氧化磷酸化作用,使病原菌无法产生所必需的能量。
唑菌酮能抑制病原菌线粒体的电子传递,主要阻断细胞色素b和细胞色素c之间的电子传递通道中ADP-ATP的氧化磷酸化作用,使病原菌无法产生所必需的能量。
类型代表品种杀菌机理抗药性风险水平苯基酰胺类甲霜灵恶霜灵苯霜灵抑制卵菌的核糖体RNA聚合酶的活性,从而抑制病菌RNA的合成,干扰病菌在寄主体内的发展作用位点单一,容易产生高水平抗药性突变菌株丙烯酰胺类烯酰吗啉氟吗啉抑制菌体内麦角甾醇的生物合成,造成细胞内物质的泄漏,最后导致菌体死亡突变频率低,突变菌株适合度差,表现为低抗药性风险甲氧基丙烯酸酯类嘧菌酯苯氧菌酯烯肟菌酯通过与细胞色素b上的辅酶QO氧化中心结合,阻止细胞ATP的合成,从而抑制线粒体的呼吸作用单一位点的突变,表现为高水平的抗药性(咪)唑类唑菌酮咪唑菌酮阻断细胞色素b和细胞色素c之间的电子传递通道中ADP-ATP的氧化磷酸化作用,抑制病原菌线粒体的电子传递田间使用较少,抗药性风险水平暂时不明确
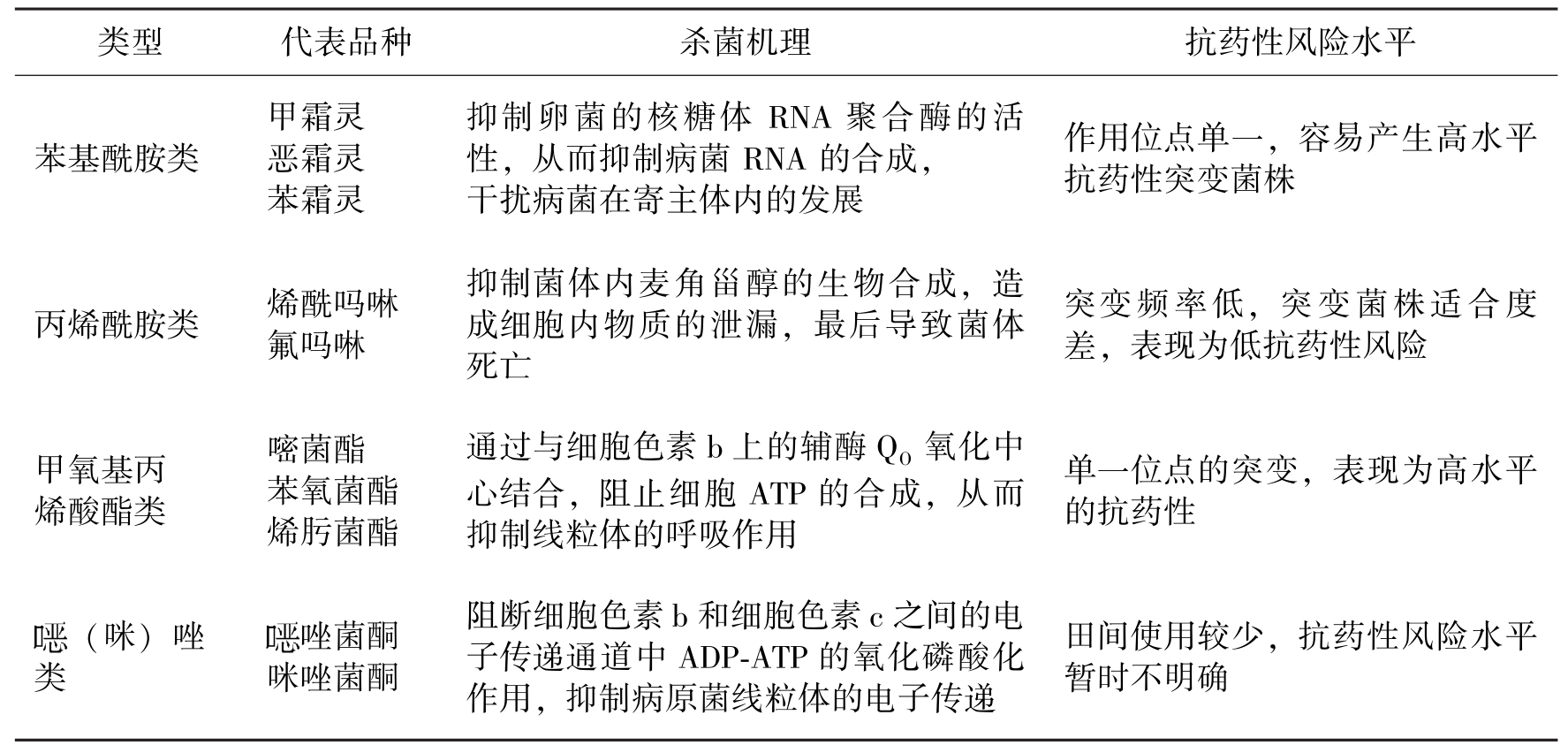
表2 防治卵菌病害的内吸性杀菌剂的主要类型、代表品种、杀菌机理、抗药性风险水平
5 结语
杀菌剂对病菌的影响是多方面的,其作用位点也相对比较复杂,由于病原菌抗药性的不断出现和抗性程度的不断上升,对研制开发广谱、新型杀菌剂的要求越来越强烈。但化学杀菌剂开发的难度越来越大,开发费用越来越高,开发新型农药的速度远远落后于病原菌抗性发展的速度。因此,明确杀菌剂的作用机理和抗性机理,对创制作用机理独特、环境友好的杀菌剂尤为重要。
卵菌的进化一般认为是从水生到陆生,由腐生到专性寄生。卵菌大部分生活在水中或潮湿的土壤中,部分比较高等接近陆生的卵菌主要寄生在高等植物体内。因此,卵菌具有一个共同的特点“亲水—疏油”。因此,在农药剂型的选择上,水剂、水乳剂、水分散粒剂、悬浮剂与卵菌有更好的亲合性,更适合卵菌病害的防治,能更好地发挥药效
因此,根据卵菌的生物学特性、卵菌病害循环的规律和药剂的作用机理,要将病害的预测预报、农药剂型、用药规范、作物种类、作物品种、当地的气候条件和当地作物的抗性水平等因素结合起来,综合考虑制定防治策略,才能保证农业的可持续发展和生态环境的和谐发展。









